


 下载:
下载:
















-
对于污水生物除磷,WENTZEL等[1]于1986年提出了强化生物除磷(enhanced biological phosphorus removal, EBPR)模型,他认为污水中的挥发性脂肪酸(volatile fatty acids, VFAs)在EBPR机制中起关键作用。研究中多以富含VFAs的废水来建立EBPR[2]。通过研究,人们获得了一些共识,即聚磷菌不能摄取水中大分子有机物来进行聚磷。若进水中只含大分子有机物,就不利于生物除磷。虽然生活污水所含的污染物主要也是大分子有机化合物如蛋白质、碳水化合物和油脂[3],但在生活污水生物处理中,由于污水中存在发酵性细菌,可将部分大分子有机化合物转化为VFAs[4],然后聚磷菌才利用这些VFAs去除污水中的磷。
1987年,TRACY等[5]在以葡萄糖为碳源的废水处理系统中也实现了EBPR,该研究发现系统内聚磷菌在胞内合成糖原而不是聚羟基脂肪酸酯。此后SATOH等[6]、ZENGIN等[7]和REY-MARTÍNEZ等[8]以葡萄糖、天冬氨酸和谷氨酸等为废水碳源,也均实现了EBPR。这些在不同碳源下的生物除磷与传统的EBPR的模型有一定差别,但相应的生化机制尚未解明。张鑫等[9]和陆雨林[10]以淀粉为唯一碳源,在缺氧/好氧序批式活性污泥法(sequencing batch reactor, SBR)与连续流系统中诱导出生物除磷,在没有厌氧释磷前提下实现了磷的去除[11],他们认为这是一种新的生物除磷机制[12]。
利用该缺氧好氧工艺,可以同时去除氮和磷,可缩短污水脱氮除磷流程、降低处理投资和运行费用。但生活污水中有机物并非单一的淀粉,而是由碳水化合物(40%左右)、蛋白质(50%左右)和油脂(10%左右)构成[3]。若进水为生活污水,对于该缺氧好氧系统的脱氮除磷情况目前尚未见相关的研究报道。本研究将与生活污水成分相近的碳源如葡萄糖、蛋白胨全脂奶粉作为补充碳源,通过设置不同的混合碳源,研究以淀粉为唯一碳源的除磷系统是否能维系以及混合碳源下生物除磷情况,旨在进一步探索该生物除磷的机理、为在生活污水生物处理中建立新型缺氧好氧生物除磷提供参考。
全文HTML
-
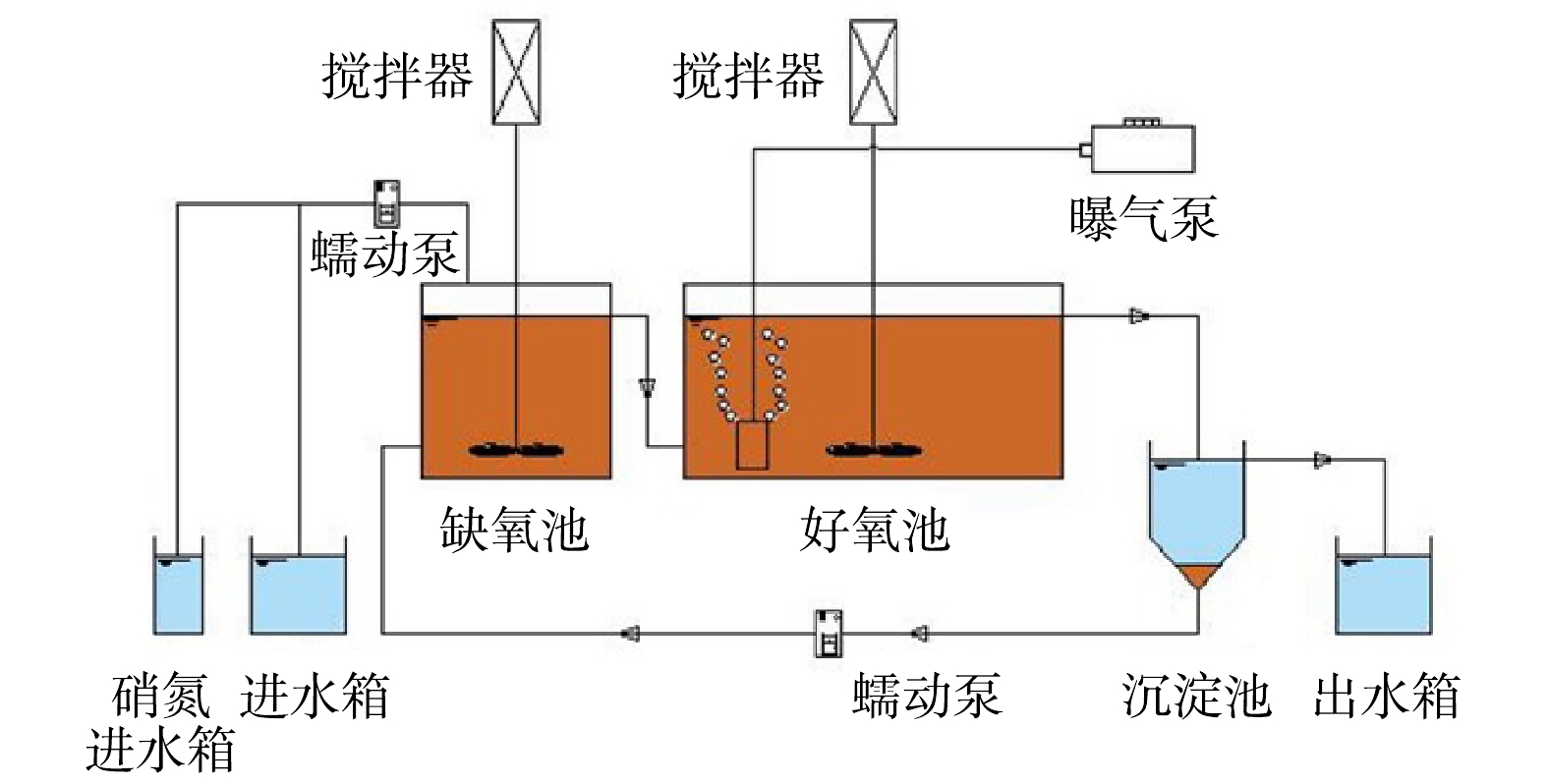
培驯污泥采用缺氧/好氧(AO)连续流系统,实验装置见图1。反应池的体积为9 L,缺氧池和好氧池体积分别为2.5 L和5.5 L,沉淀池体积为2.4 L,反应器的温度为室温,进水流量为24 L·d−1,水力停留时间为9 h。缺氧池进行机械搅拌,好氧池溶解氧(dissolved oxygen, DO)控制在0.5~1.0 mg·L−1。每天从好氧池末端排出污泥来控制污泥停留时间为20 d左右,维持污泥浓度2 800~3 400 mg·L−1。进水中氮仅为硝酸盐态氮,在缺氧与好氧段之间不设内回流。二沉池污泥回流缺氧池,回流比为100%。
-
本实验接种污泥取自西安某污水处理厂,培驯污泥进水采用人工废水,以可溶性淀粉为碳源(COD约为400 mg·L−1),磷酸二氢钾(
PO3−4 -P浓度约为8 mg·L−1)和硝酸钾(NO−3 -N浓度约为20 mg·L−1),再加入硫酸镁0.1 g·L−1、硫化钙0.01 g·L−1以维持微生物良好生长。 -
COD、
NO−3 -N、PO3−4 -P等常规指标的测定采用文献中的方法[13]。乳酸的测定采用对羟基联苯法[14],糖原采用蒽酮比色法[15],PHB预处理后采用气相色谱法(Agilent 6890N,FID检测器,HP-5型色谱柱)测定,进样方式为自动进样。色谱条件为分流比2∶1,进样口温度和检测器温度分别为250 ℃和300 ℃;采用柱箱升温,起始炉温为50 ℃,保留1 min,以10 ℃·min−1 升到120 ℃,再120 ℃·min−1 升到270 ℃,保留6 min,进样量为2 μL[16]。污泥中聚磷颗粒染色方法采用4',6-二咪基-2-苯基吲哚(4',6-diamidino-2-phenylindole dihydrochloride, DAPI)溶液避光染色[17]。 -
从反应器[18]好氧段取1 L活性污泥,在转速为4 000 r·min−1下离心5 min后,弃去上清液后,加蒸馏水重复操作上述步骤3次,再用蒸馏水将沉淀污泥定容至500 mL,并加入500 mL人工模拟废水,配置总COD为400 mg·L−1的复合碳源组成废水(表1),探究复合碳源中淀粉浓度对除磷的影响。固定淀粉浓度为400 mg·L−1,配制总COD分别为500 mg·L−1和600 mg·L−1的复合碳源废水(表2),探究葡萄糖碳源以及蛋白类碳源对除磷的影响。批式实验总时长为9 h,缺氧段为2.5 h,好氧段为6.5 h,从添加人工模拟废水开始计时并取第1个样,而后缺氧阶段隔30 min取样,好氧阶段每隔60 min取样,测定
PO3−4 -P、乳酸和糖原的含量。连续流缺氧段除磷物料平衡按式(1)进行计算。式中:R为磷酸盐去除率;PA-inf和 PA-eff分别为缺氧池进出水
PO3−4 -P浓度,mg·L−1;VSS,A为缺氧池VSS浓度,mg·L−1;TA为缺氧池反应时间,h。
1.1. 实验装置
1.2. 培驯污泥阶段进水水质
1.3. 分析方法
1.4. 实验方案
-
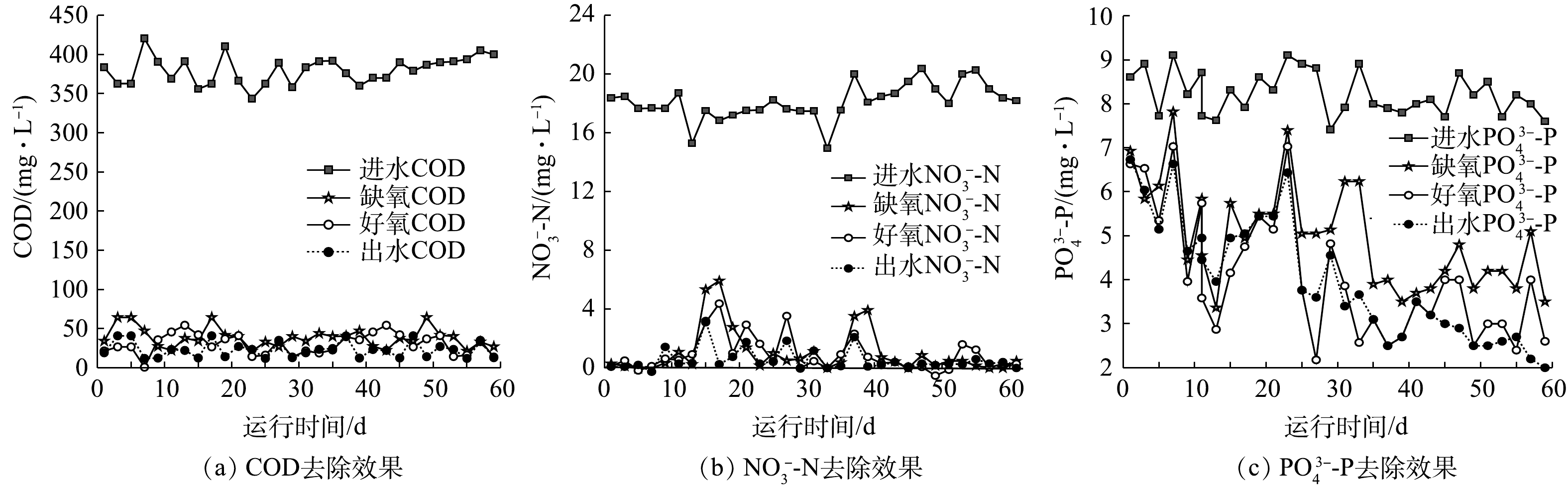
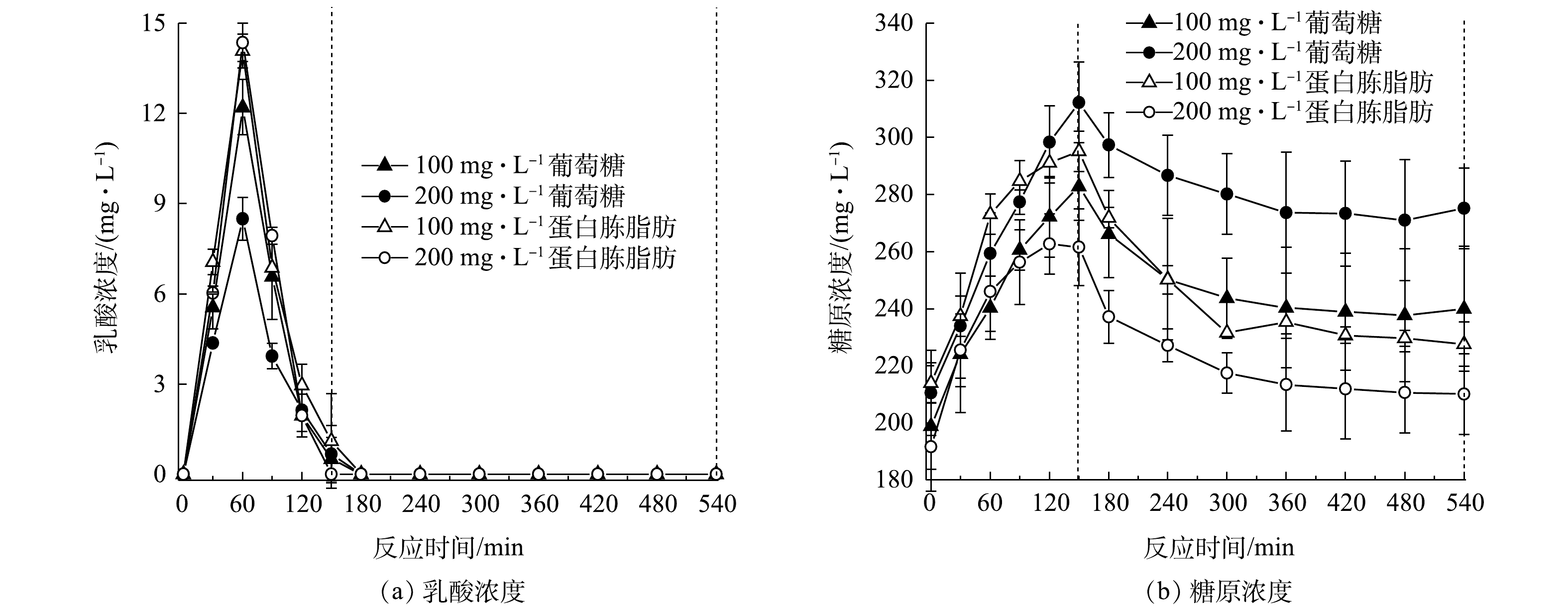
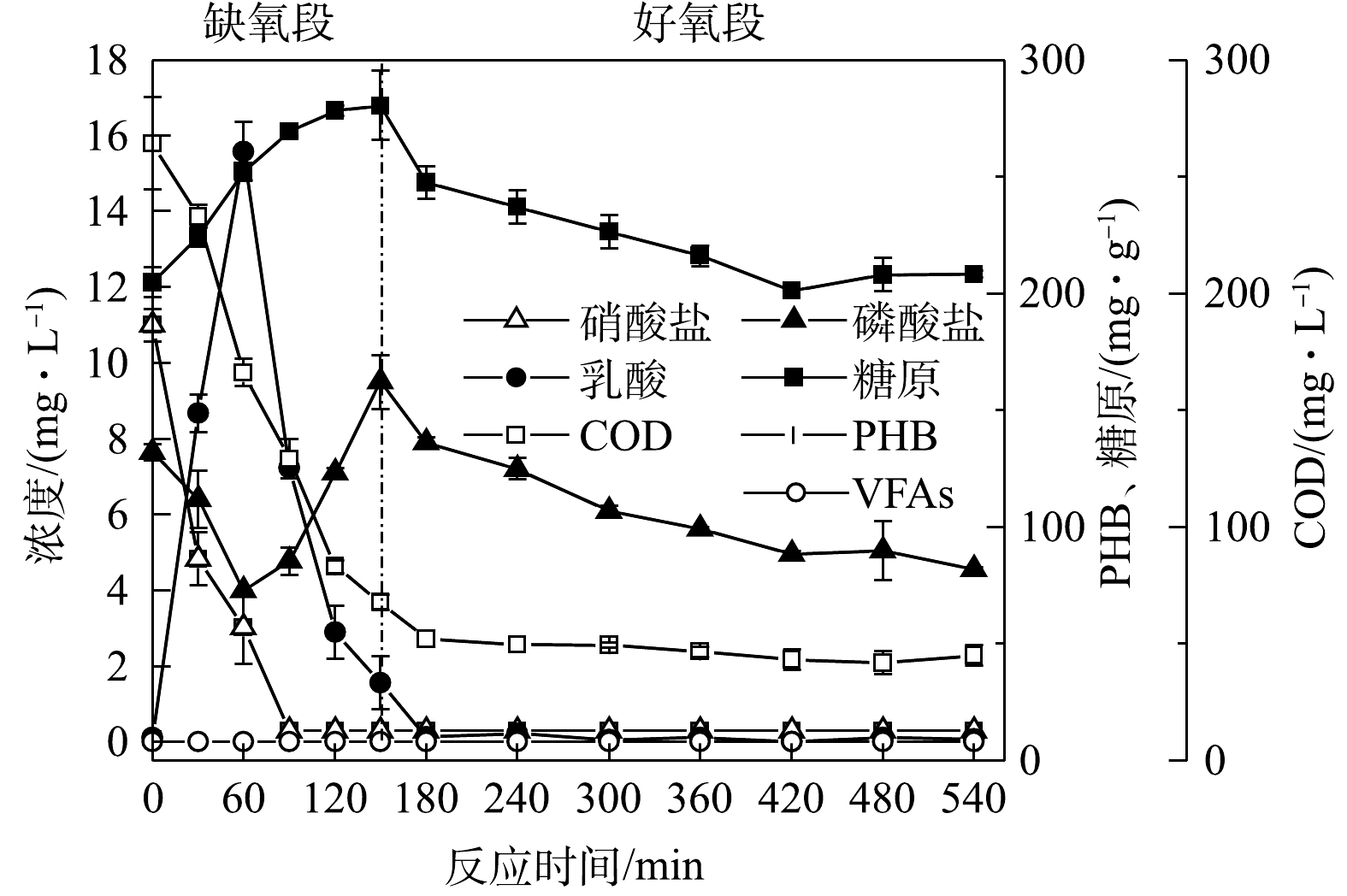
图2为连续流系统对废水中污染物的去除结果。由图2可见,反应器经过40 d的驯化后,除磷效果趋于稳定,出水
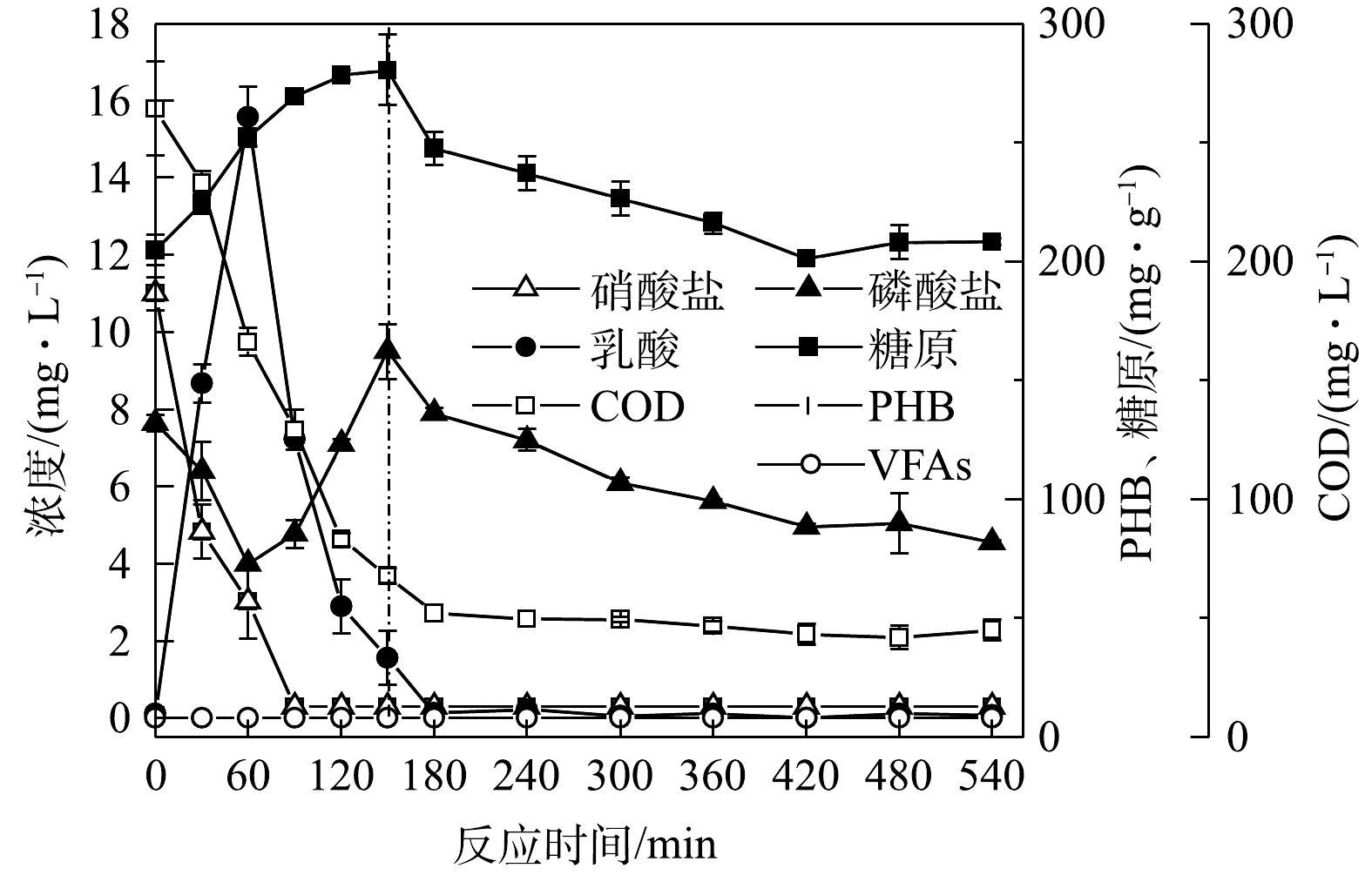
PO3−4 -P为2.6 mg·L−1,去除率达到72%左右,缺氧段除磷量为3.5~4 mg·L−1,去除率达到50%;出水COD小于50 mg·L−1,去除率达95.6%;出水的NO−3 -N为0.3 mg·L−1,去除率达98.6%,全程没有释放磷的现象产生。JIN等[19]的研究也表明,磷的去除可以在厌氧不释放磷的情况下实现。系统在无厌氧条件下,在缺氧段与好氧段均可实现磷去除,故其为新型生物除磷。为研究该新型缺氧好氧除磷特征,取系统污泥进行批式实验,结果见图3。由图3可知,在缺氧段,糖原含量由 196.78 mg·g−1 (以VSS计)增至280.44 mg·g−1,缺氧段乳酸含量达到15.6 mg·L−1,水中的磷酸盐浓度随之降低,缺氧段除磷为3.3 mg·L−1,
PO3−4 -P去除率在40%左右。这说明在缺氧段,除磷的能量为产酸菌在糖酵解产乳酸过程中生成的ATP[12]。在好氧段,糖原含量由280.44 mg·g−1降低到208.26 mg·g−1,好氧段除磷为3.2 mg·L−1。在传统生物除磷中,聚磷菌(polyphosphate accumulating bacteria, PAOs)将VFAs以PHA的形式储存,并在厌氧段充分释磷。在好氧阶段,PHA被用作生长和补充体内糖原和多聚磷库的碳源和能量来源[20]。该系统(图3)中未检测到聚β羟基丁酸(polyhydroxybutyric acid, PHB)和VFAs,碳源在缺氧段以糖原的形式累积,这与王琰[21]的研究结果一致。这说明在好氧阶段中聚磷菌吸磷的能量主要来自糖原的氧化。
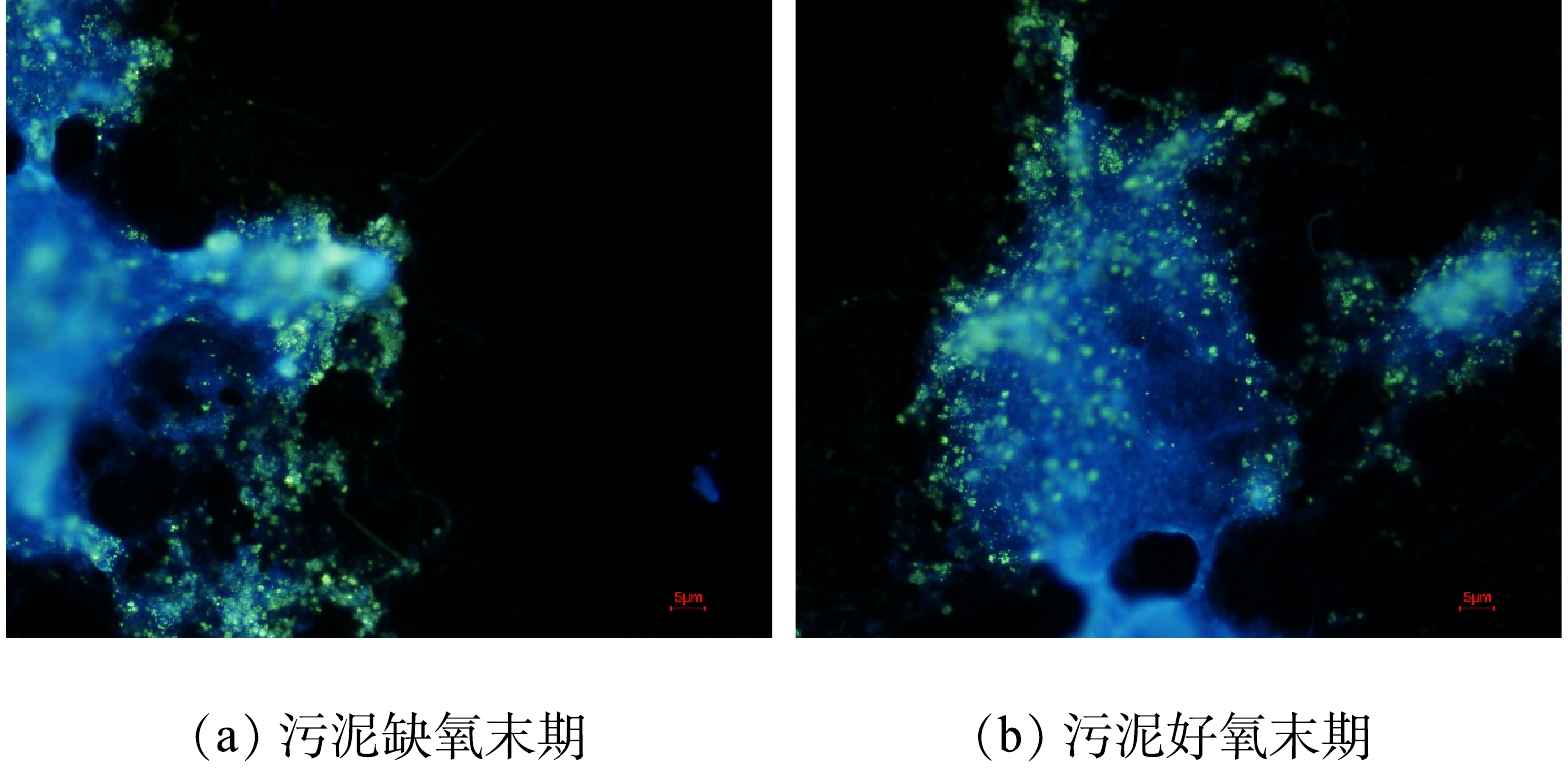
由于批式实验为一次性进水,在缺氧后期缺少硝酸盐,进入了厌氧环境,PAOs出现释磷现象,这与新型缺氧好氧除磷连续流系统存在差异。为解释批式实验与连续流系统存在的矛盾,对AO连续流污泥聚磷颗粒进行了DAPI染色,镜检照片如图4所示。可见细菌的细胞呈蓝色,聚磷颗粒呈亮黄色颗粒。连续流缺氧段(图4(a)和好氧段(图4(b))聚磷颗粒分布以及数量差异较小,好氧末聚磷颗粒较缺氧末略微增多,好氧末污泥含磷率较缺氧末轻微增长,这说明系统除磷主要集中在缺氧阶段,同时系统中微生物表现为单一的吸磷过程。
在新型缺氧好氧除磷系统中,缺氧阶段存在聚磷和释磷2种现象,系统中新型的缺氧除磷菌为优势菌种,因此,缺氧吸磷大于释磷。在好氧段,PAOs利用胞内的糖原氧化可以进一步摄取磷。故而AO连续流的缺氧阶段总体上是净摄取水中磷。这一结果与霍小爱等[11]和LUO等[12]报道的在AO-SBR中除磷过程以及传统厌氧好氧系统中的生物除磷过程均不同。
-
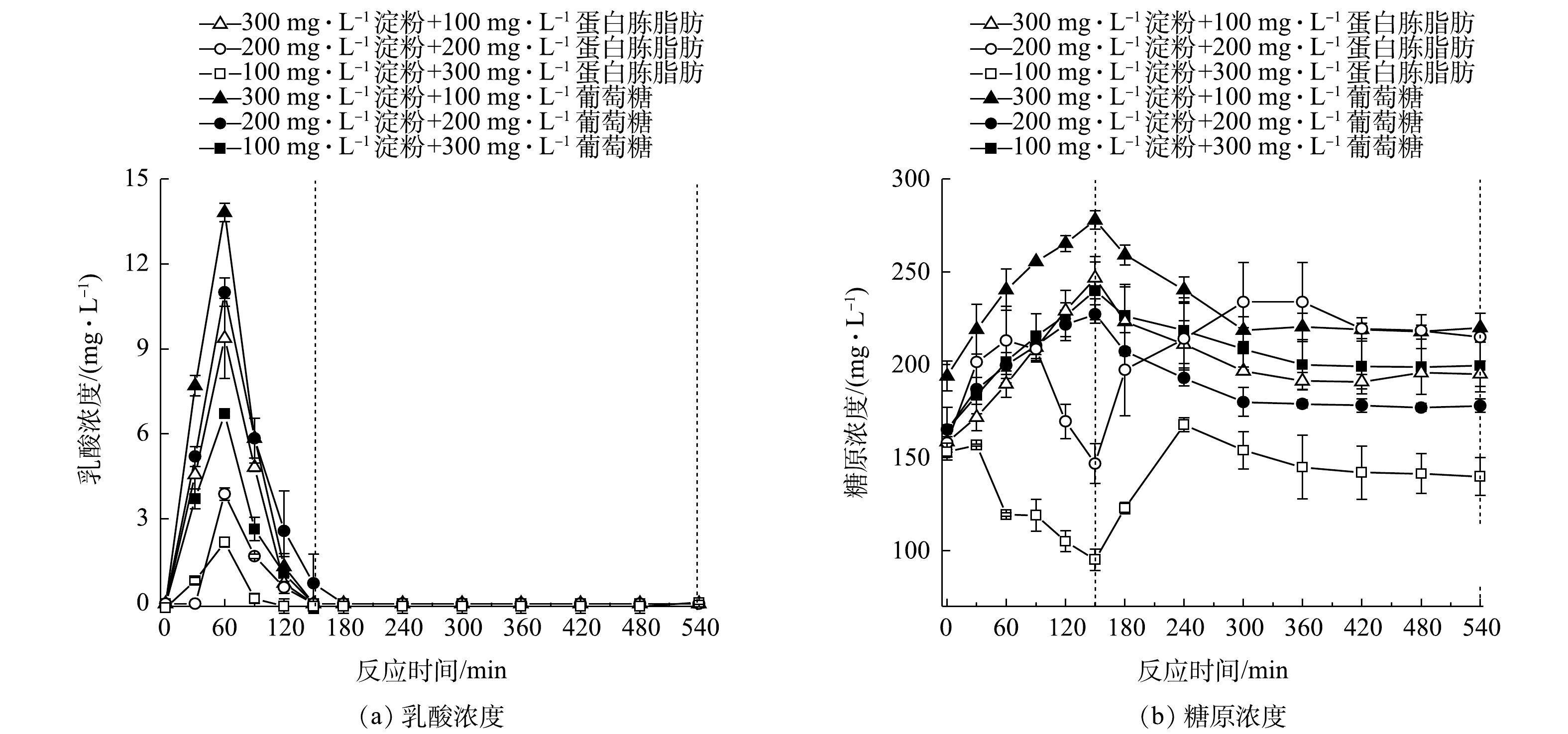
为探究复合碳源(葡萄糖与蛋白类碳源)对系统除磷菌活性的影响,保持淀粉含量(以COD计)约为400 mg·L−1,加入蛋白胨全脂奶粉、葡萄糖配制总COD为500 mg·L−1和600 mg·L−1的复合碳源废水进行批式实验,系统的除磷效果如图5所示。由图5可见,加入葡萄糖配制总COD为500 mg·L−1和600 mg·L−1的复合碳源的废水后,缺氧段除磷分别为2.2 mg·L−1和0.8 mg·L−1,污泥的吸磷速率分别为0.71 mg·(g·h)−1和0.27 mg·(g·h)−1;好氧段除磷分别为2.4 mg·L−1和1.8 mg·L−1。加入蛋白胨全脂奶粉配制总COD为500 mg·L−1和600 mg·L−1的复合碳源废水后,系统缺氧段除磷分别为2.9 mg·L−1和2.6 mg·L−1,污泥吸磷速率分别为0.93 mg·(g·h)−1和0.89 mg·(g·h)−1,好氧段对磷的去除分别可达3.4 mg·L−1和3.0 mg·L−1。
缺氧段系统液相中乳酸含量以及污泥中糖原量变化如图6(a)和图6(b)所示。由图6可见,供给加入葡萄糖总COD为500 mg·L−1和600 mg·L−1的复合碳源废水后,缺氧段水中乳酸含量分别为12.2 mg·L−1和8.5 mg·L−1;污泥中糖原含量分别为282.9 mg·g−1和312.3 mg·g−1;好氧段糖原消耗量分别为43.0 mg·g−1和37.1 mg·g−1。供给加入蛋白胨全脂奶粉总COD为500 mg·L−1和600 mg·L−1的复合碳源废水后,缺氧段水中乳酸量分别为14.1 mg·L−1和4.4 mg·L−1,好氧段糖原消耗量分别为67.34 mg·g−1和52.57 mg·g−1。
上述结果说明,当进水中有充足的淀粉时,再投加葡萄糖,过量碳源就会抑制新型缺氧好氧除磷[22],磷的去除量会有所下降。在不同复合碳源组成下,缺氧阶段糖原大量合成,而好氧段糖原消耗却减少,说明糖原没有被聚磷菌储存,新型缺氧好氧除磷系统中可能存在一定量的聚糖菌(glycogen accumulating organisms, GAO)[23],其能够在缺氧条件下利用碳源而不释放磷酸盐,糖原在没有能量消耗的情况下合成,聚糖菌在底物竞争中取得优势,导致产酸量减少,从而使得除磷效果变差。因此,在碳源充足时,投加葡萄糖会刺激聚糖菌代谢,抑制新型缺氧好氧除磷除磷。
对于传统生物除磷,一般水中有机物越充足,则除磷的效果会越好[24]。但在本研究中,当水中淀粉含量充足、再多加入些蛋白胨脂肪后,除磷效果也并未提高,且缺氧段也未见有更多的乳酸生成(图6(a)),这从侧面说明蛋白类碳源对依靠淀粉发酵产乳酸来进行的生物除磷是不起作用的。
-
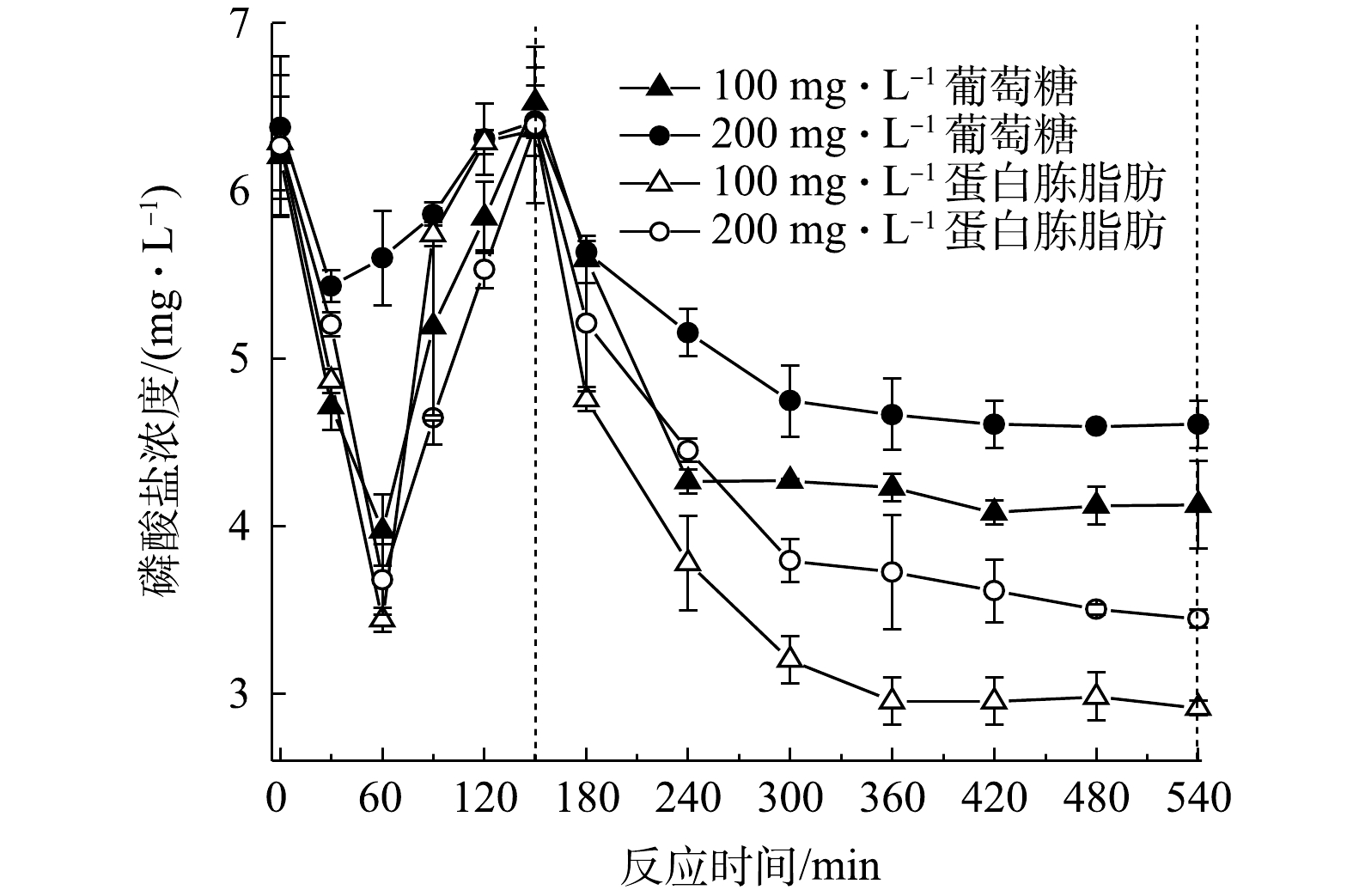
保持总COD为 400 mg·L−1不变,将进水中的淀粉量减少至300、200、100 mg·L−1,并相应增加葡萄糖或蛋白胨与全脂奶粉混合物进行批式实验,系统除磷效果如图7所示。由图7可见,当葡萄糖分别为100、200、300 mg·L−1时,系统缺氧段除磷分别为2.9、2.4和0.6 mg·L−1,好氧段除磷分别为3.3、2.1、1.9 mg·L−1。当含蛋白胨全脂奶粉浓度为100 mg·L−1下,缺氧段除磷为2.2 mg·L−1,好氧段除磷为2.9 mg·L−1;当含蛋白胨全脂奶粉浓度为200 mg·L−1和300 mg·L−1下,系统不具备新型缺氧好氧除磷特征,为传统厌氧释磷好氧除磷特征。
在不同复合碳源下的缺氧段乳酸产生量和污泥中糖原含量变化结果见图8(a)和图8(b)。当进水葡萄糖含量为100、200、300 mg·L−1时,缺氧段乳酸的量分别为13.8、11.1、6.7 mg·L−1,好氧段糖原的消耗量分别为57.9、49.2、40.2 mg·g−1。当进水蛋白胨全脂奶粉浓度为100 mg·L−1下时,缺氧段乳酸量为9.4 mg·L−1,好氧段糖原的消耗量为51.64 mg·g−1;当进水含蛋白胨全脂奶粉量为200 mg·L−1和300 mg·L−1下,缺氧段乳酸量分别为3.9 mg·L−1和2.1 mg·L−1,好氧段糖原不断累积,这与传统PAOs的糖原代谢方式相同。
结合图7和图8可看出,进水含蛋白胨全脂奶粉与含淀粉混合碳源下的系统除磷效果较差。其原因可能是,尽管系统中新型缺氧好氧除磷污泥具有专性发酵功能[25],但其不能有效利用蛋白胨和全脂奶粉进行发酵产乳酸,乳酸量减少,则相关细菌产生可用于摄磷的ATP减少,进而影响到除磷效果。
由此可见,该新型缺氧好氧除磷系统中淀粉浓度是影响除磷效果的关键。降低淀粉浓度,会导致缺氧阶段乳酸浓度减少,从而影响除磷效果。当葡萄糖浓度为100 mg·L−1淀粉为300 mg·L−1时,缺氧段乳酸产生量最多,糖原在好氧段消耗量也最大,除磷效果最优。因此,在以淀粉为唯一碳源的系统中,葡萄糖可在淀粉不充足时补充碳源用于除磷。
2.1. AO连续流系统运行性能
2.2. 进水变为复合碳源、COD大于400 mg·L−1对污泥除磷的影响
2.3. 进水总COD为400 mg·L−1下复合碳源对污泥除磷的影响
-
1)在以淀粉为碳源下,AO连续流生物脱氮系统缺氧段除磷为产酸菌通过淀粉发酵产乳酸提供能量进行超量摄磷,在好氧段系统中存在的传统PAOs通过糖原提供能量进行摄磷,在这2类除磷菌作用下,实现了好氧缺氧条件下的废水中磷的去除。
2)进水中可溶性淀粉的量减少,会导致乳酸的产量减少和除磷效果变差。
3)缺氧好氧除磷污泥不能利用蛋白类碳源进行除磷代谢,系统内发酵产酸菌能够代谢葡萄糖并产乳酸用于除磷,一定的葡萄糖可以弥补淀粉不足对除磷的影响,但淀粉充足时,葡萄糖或者蛋白类碳源的存在对除磷会不利。故该生物除磷还是很难在含多种碳源的污水中实现高效除磷。
