

























-
随着工业化和城市化的推进,有机固体废物产量逐年增加[1]。这些固体废物中有机组分含量高,可以通过厌氧发酵生产挥发性脂肪酸(VFA)[2]。但VFA几乎完全溶于水,难以分离和提纯[3]。脂肪酸的水溶性随着碳链的延长而逐渐降低,其中碳链大于6的中链脂肪酸(MCFA)具有较好的疏水性。例如:正丁酸与水互溶,正己酸的溶解度则仅为10.82 g·L−1,正辛酸的溶解度则几乎为0[4]。因此,将VFA加长碳链后,其产物的后续分离纯化将更容易。例如,MCFA可以通过离子交换树脂提取[1]、膜萃取[5-6]等方式进行分离。与VFA相比,MCFA除更易分离外,其附加值也更高。有研究[7]表明,乙酸(工业级)的价格约为3 000元·t−1,而己酸(工业级)的价格高达1.7×104 元·t−1。同时,MCFA还是生物柴油、生物塑料、抗菌剂和香料等的前驱体[8],具有较高的应用价值。综上所述,利用微生物链延长技术将VFA合成为MCFA,已得到越来越多的关注。
目前,已经报道的可将VFA链延长从而合成MCFA的微生物有5种。其中,克氏梭菌Clostridium kluyveri由于pH适应范围较广(pH为5.2~8.0),且具有产孢子、耐高温等特性,故研究最多、应用最广[6]。通常,克氏梭菌利用乙醇、乳酸、氢气作为电子供体,将乙酸、丙酸、丁酸等短链羧酸作为电子受体,通过反向β氧化途径生成MCFA。在反向β氧化途径中,碳链每次仅延长2个碳原子,例如丁酸(C4)是由乙酸(C2)和乙醇链延长而成的,己酸(C6)则是由丁酸(C4)和乙醇链延长而成的[6]。而且,反向β氧化途径是一个循环过程,在这个循环过程中,乙醇被NAD+氧化生成乙酰辅酶A,引起底物水平磷酸化,H+在铁氧化还原蛋白作用下被还原为H2,产生膜内外质子浓度差[9]。
由于生物质厌氧发酵液中含有大量的VFA,因此,微生物链延长产MCFA工艺可以以易降解生物质的厌氧发酵液作为底物。例如,污泥、餐厨垃圾、果蔬垃圾和木质纤维素类有机废物均可以经过厌氧发酵产生富含VFA的厌氧发酵液,然后将此厌氧发酵液投加到链延长工艺系统中合成MCFA[10]。生产出的MCFA可以作为医药生产中的抗菌剂、食品行业中的风味添加剂、家禽饲料和猪饲料中的添加剂、化工生产原料、生物柴油的前驱体等[9,11]。总之,利用有机废物的厌氧发酵液进行微生物链延长,一方面可以实现有机废物的资源化利用,另一方面可以生产高值产品MCFA[9]。这使得以链延长为基础的生物法已经成为MCFA生产的一种新兴方法[4]。
本文以微生物链延长生产MCFA为中心,梳理了有机废物厌氧发酵液作为底物的微生物链延长机理和主要影响因素,总结了单一有机废物厌氧发酵液和混合有机废物厌氧发酵液作为底物的微生物链延长技术研究进展,并提出了今后有机废物厌氧发酵液链延长技术的研究方向,以期为生物质的资源化和微生物链延长技术研究提供参考。
-
链延长过程涉及电子供体的氧化和反向β氧化过程[12]。如图1所示,反向β氧化过程是电子供体(乙醇或乳酸)与电子受体(短链脂肪酸)链延长生成MCFA代谢途径中的核心过程,该过程由电子供体提供碳源、能量和还原当量[9]。首先,乙醇被氧化为乙醛,在乙醇脱氢酶和乙醛脱氢酶的催化作用下生成乙酰辅酶A。其中,大约有1/6的乙酰辅酶A通过底物水平磷酸化转化为乙酸,并释放能量;另外5/6的乙酰辅酶A进入反向β氧化循环[9]。在乙酸链延长生成正丁酸的第一个循环(RBO 1)中,乙酰辅酶A与另一个乙酰辅酶A在乙酰辅酶A硫代酶的作用下偶联,形成乙酰乙酰辅酶A,之后经过一系列酶促反应,生成关键中间产物丁酰辅酶A[4];在乙酰辅酶A转移酶的作用下,一个丁酰辅酶A转移至电子受体乙酸,生成正丁酸并释放乙酰辅酶A;释放的乙酰辅酶A与乙醇氧化生成的乙酰辅酶A反应,再次进入RBO 1中继续乙酸的链延长[13]。类似地,在正丁酸链延长生成正己酸的第二个循环(RBO 2)中,经过一系列酶促反应,乙酰辅酶A与丁酰辅酶A结合,生成关键中间产物己酰辅酶A,己酰辅酶A转移至丁酸,生成正己酸并释放丁酰辅酶A,释放的丁酰辅酶A与乙醇氧化生成的乙酰辅酶A反应,再次进入RBO 2中继续正丁酸的链延长[2]。在乙醇或乳酸为电子供体的链延长反应中,乙酸可以被顺序链延长为丁酸、己酸和辛酸,丙酸可以被顺序链延长为戊酸、庚酸和壬酸。
有机废物厌氧发酵液中包括乙酸、丙酸、正丁酸等,一般乙酸浓度最高[11]。以生成正己酸为例,基于克氏梭菌链延长合成代谢和分解代谢的路径如图2所示。有机废物厌氧发酵液的链延长过程主要包括正己酸合成(R1)和正戊酸合成(R2);竞争过程主要包括乙醇氧化(R3)、乙酸氧化(R4)、丙酸氧化(R5)、正丁酸氧化(R6)、甲烷生成(R7)等。在微生物链延长过程中,当底物为纯物质乙酸和乙醇时,乙酸先经过链延长生成正丁酸,然后正丁酸继续链延长生成正己酸[11,14]。LIU等[14]将乙醇和乙酸按照3∶1的质量比加入有效容积为1 L的链延长系统中,达到的最大己酸质量浓度为3.03 g·L−1。YIN等[15]按照7∶3的比例(质量比)向有效容积100 mL的链延长系统中添加乙醇和乙酸,生成的己酸的质量浓度为3.11 g·L−1。BAO等[16]将乙酸、丙酸、丁酸分别和乙醇按照不同比例在100 mL有效容积的序批式反应器中进行微生物链延长合成己酸,得到的最高己酸的质量浓度分别为2.33、2.49和4.82 g·L−1,即丁酸的微生物链延长效率更高。有机废物厌氧发酵液微生物链延长合成己酸的主要代谢反应如表1所示。由于有机废物厌氧发酵液中含有正丁酸[11],可以直接与乙醇进行微生物链延长合成正己酸,故相比于纯物质乙酸和乙醇链延长工艺生产正己酸,厌氧发酵液作为底物的链延长工艺可以提高己酸产率,同时减少乙醇的消耗,并实现有机废物的高效资源化。包率[17]将乙酸、丙酸、丁酸按一定比例配成混合酸,投加到100 mL有效容积的链延长工艺系统中,生成的己酸浓度达到6.58 g·L−1,这表明混合酸微生物链延长比单酸作为底物可以生成更高产量的己酸。因此,利用含有乙酸、丙酸、丁酸等的有机废物厌氧发酵液进行链延长,可以得到更高浓度的MCFA。
-
克氏梭菌适应的温度范围较广,一般在19~45 ℃条件下均可以进行链延长。AGLER等[18]在容积为5 L的厌氧序批式反应器中进行链延长实验,对比了温度为55和30 ℃时的链延长效果;结果表明,当温度为55 ℃时,克氏梭菌未能促进己酸的合成,而将温度降低至30 ℃时,则己酸产量不断增加。在目前的研究中,通常将链延长实验的温度控制在30~40 ℃。大多数的链延长实验在30 ℃时得到较高的己酸产量和产率[19-20],这可能由于MCFA的溶解性增加了细胞膜的流动性,可以在比克氏梭菌最佳生长速率(34 ℃)更低的温度下获得MCFA的最大产率[19]。
pH是反映厌氧发酵链延长系统稳定性的重要指标之一。pH能够显著影响微生物的新陈代谢以及酶的活性;同时,体系pH也受微生物的生长代谢过程的影响[21]。当系统pH处于6.75~7.00时,厌氧发酵液微生物链延长的己酸产量最高。这主要是因为,如果非解离态己酸的质量浓度过高(超过0.79 g·L−1)时可抑制克氏梭菌的生命活动。而在弱酸性条件下,链延长产生的非解离态己酸较少,毒性较小,因此,中性偏弱酸性的环境(pH=6.75~7.00)有利于微生物链延长反应的进行[11,16]。WEIMER等[22]通过研究pH(4.8~9.2)对3231B梭状芽孢杆菌生长的影响,发现pH为6.8时己酸质量浓度达到最高,为12.8 g·L−1。STEINBUSCH等[23]在125 mL有效容积的血清瓶中用乙酸和乙醇作为底物分别在pH为5.5和7.0条件下进行链延长实验;结果表明,在pH=7.0时,己酸的质量浓度和产率分别为8.27 g·L−1和25.6 g·(L·d)−1,比pH=5.5时分别高8.15 g·L−1和21.7 g·(L·d)−1。虽然链延长系统的初始pH容易控制,但在反应进行过程中,pH会不断下降[4,11,17]。 pH的降低可直接影响克氏梭菌的生命活动。例如,LIU等[24]用乙醇和乙酸进行链延长实验,将初始pH调至中性区间;结果表明,随着链延长反应的进行,pH不断降低,当pH在第20 d降至5.5以下时,己酸产率开始降低,当加入氢氧化钾使pH恢复至中性区间后,己酸产率又大幅度提高。pH还可以从不同方面间接影响己酸的合成,如pH降低可以促使乙酸还原为乙醇,故不利于己酸的合成[4]。因此,pH过低不利于微生物链延长产己酸反应。
-
有研究者发现,厌氧发酵液链延长所产己酸的浓度与有机废物厌氧发酵过程中的底物浓度(含固率)密切相关[11]。通常,有机废物浓度越高,厌氧发酵液中的VFA浓度相应越高,在后续微生物链延长过程中己酸的产量亦越高。但当厌氧发酵液中乙酸和正丁酸的质量浓度过高,分别超过7.9和19.4 g·L−1时,可对链延长生产己酸反应具有抑制作用[25]。WANG等[11]研究了300 mL序批式反应器中污泥含固率(2%~8%)对水解酸化液链延长的影响;结果表明,在8%含固率的污泥水解酸化液中VFA浓度最高,且各种酸的比例更适合微生物链延长反应,链延长获得的己酸质量浓度达到5.04 g·L−1。BOLAJI等[26]用含固率为4.82%的果蔬废物为底物,在300 mL的半连续式反应器(有效容积150 mL)中厌氧发酵后进行链延长实验,发现发酵液中同时存在短链脂肪酸和中链脂肪酸;其中,丁酸是主要发酵产物,其次是己酸,己酸的质量浓度最高达到8.78 g·L−1。YU等[27]利用含固率为14.4%的果蔬废物作为底物,在有效容积为1 L的反应器中进行两相连续式微生物链延长实验,在水解酸化相中,接种物与底物的质量比为2∶1,厌氧发酵液中乙醇、乙酸、丁酸的质量浓度分别为0.8、5.3、3.6 g·L−1,产生的水解酸化液流入第二相后,按照乙醇/酸(质量比)为4∶1的比例将乙醇投加到链延长系统中,己酸的质量浓度可达14.9 g·L−1。
-
水力停留时间(HRT)和污泥停留时间(SRT)对于控制厌氧过程至关重要,这是因为它们与反应器中活性微生物的数量和质量、以及生物相的组成有直接关系[9]。降低反应器HRT或SRT可促进链延长过程,同时抑制产甲烷过程。降低HRT,一方面产甲烷菌可以在达到对数期之前从反应器中排出,无法在反应器中积累;另一方面乙酸型产甲烷菌的生长可被抑制,从而有效抑制产甲烷,减少乙酸的消耗[4]。同时,缩短HRT可使反应器中产生的己酸及时排出,减少其对系统中链延长微生物的毒性,从而提高系统的己酸合成速率[9]。GROOTSCHOLTEN等[19]将1 L反应器的HRT从16 h缩短至4 h,产物中己酸的质量浓度由12.0 g·L−1降低至9.3 g·L−1,但己酸合成速率由16.0 g·(L·d)−1提高到55.8 g·(L·d)−1。这主要是由于HRT的降低限制了乙酸型产甲烷菌的活性。虽然HRT缩短可使己酸浓度降低,但己酸生产的选择率高于80%[4]。因此,在完全搅拌且生物量分散均匀的反应器中,相应减少HRT对产己酸有利。在单相固体废物厌氧发酵链延长过程中,控制HRT是可行的,而SRT的改变可以通过控制HRT来实现[9]。
-
在不同条件下,利用不同底物、不同反应器进行链延长生产己酸的效果比较如表2所示。H2可以作为链延长的电子供体,因此,氢分压对链延长合成己酸反应有显著影响[17]。有研究表明,在低氢分压条件下,底物短链羧酸和产物MCFA均可通过β氧化途径被降解;相反,高氢分压可以在乙醇消耗减少时弥补还原当量的不足,继续与电子受体反应生成己酸,同时抑制乙醇、短链羧酸和MCFA的厌氧氧化,促进短链羧酸向MCFA产物方向进行链延长[8,17]。
在链延长过程中,克氏梭菌通常以乙醇作为电子供体进行己酸的生产;同时,克氏梭菌在合成代谢过程中对无机碳源(CO2)也有一定需求[9]。有研究表明,在克氏梭菌生产己酸过程中,微生物生长吸收的碳约30%来自于CO2,其余来自有机碳源[4,17]。由于克氏梭菌利用CO2合成氨基酸的速率高于利用HCO3−的速率,因此,通常认为克氏梭菌的生长速率与其所利用的CO2浓度密切相关[4]。当链延长底物为乙醇和乙酸时,通常需要外加CO2提供无机碳源。当在两相厌氧发酵链延长反应器中以复杂有机物为底物时,这些复杂有机物在初级厌氧发酵时可产生内源CO2,并进入第二相参与链延长微生物的生长繁殖,从而促进己酸合成。但当体系中产生的CO2过剩时,体系的酸度有所提升,产甲烷菌的活性被抑制,同时系统氢分压也随之降低[4],促进VFA的氧化,不利于己酸合成[8]。因此,采用厌氧发酵液作为底物进行链延长通常不需要补充无机碳源。
-
电子供体在链延长过程中发挥着重要作用,可提供链延长过程中必不可少的碳源、能源和还原当量[5]。可以作为电子供体的物质很多,如乙醇、乳酸、H2和CO等[9]。电子供体类型不同,链延长反应的机理也不同。目前,大多数研究以乙醇和乳酸作为链延长反应的电子供体[32]。
电子供体的质量浓度对链延长反应具有较大影响,其中许多研究关注乙醇投加量对链延长反应的影响。LIU等[14]探究了不同乙醇/乙酸质量比对正己酸产量的影响,发现乙醇/乙酸质量比大于2∶1时,才能保证链延长反应有足够的电子供体;当乙醇/乙酸质量比为3∶1时,链延长生成的正己酸的质量浓度最高,为3.03 g·L−1。但是,高浓度的乙醇(200~400 mmol·L−1)可对链延长菌产生毒害作用[6]。LONKAR等[28]发现,当乙醇质量浓度为10 g·L−1时,生成的MCFA质量浓度可以高达10 g·L−1,但当乙醇质量浓度高于40 g·L−1时,不在发生链延长反应。乙醇的毒性可以通过稀释底物浓度的方法进行削减,但该方法也可导致MCFA产量降低。在自然界中,乳酸是较易获得的电子供体,故近年来针对乳酸链延长的研究也在增多[9]。KUCEK等[33]在一个连续流反应器中将乳酸转化为正己酸,最高正己酸产率为3.12 g·(L·d)−1。乳酸浓度对链延长反应的影响较大,高浓度乳酸会促使乳酸中的碳通过竞争途径转化为丙酸[34]。尽管低浓度乳酸亦可促进链延长过程,但由于其电子供体浓度较低,故得到的MCFA产量不高[34]。未来的研究可关注解除乳酸浓度抑制、提高链延长效率的方法。
-
富含厌氧微生物菌群的开放培养体系能够有效地将复杂有机废物转化为特定的生物化学产品。与无菌系统相比,开放培养体系的运行成本相对更低[6]。克氏梭菌是链延长反应中的关键微生物,可利用乙醇、乳酸、H2等作为电子供体,将乙酸等短链羧酸作为电子受体,通过反向β氧化途径生产MCFA [35]。
利用克氏梭菌进行链延长反应有2种方式。一种方式是直接加入以乙醇(或乳酸)和乙酸为底物驯化富集的链延长污泥。WU等[1]在有效容积为10 L的反应器中加入质量浓度为1.98 g·L−1活性污泥碱性发酵液进行链延长实验;结果表明,加入4.8 g驯化的链延长菌,链延长反应的迟滞期降低为2 d,MCFA中碳链较长的正辛酸占比最高,当乙醇投加量为3.46 g·L−1时,达到最高中链脂肪酸产率1.49 g·(L·d)−1。另一种方式,在链延长反应前,先经过一个污泥驯化阶段,在反应器中富集较高丰度的克氏梭菌,再投加底物进行链延长反应。GROOTSCHOLTEN等[7]利用城市有机固体废物进行厌氧发酵实验,发酵液VFA质量浓度为6.9 g·L−1,然后将发酵液作为底物加入1 L反应器中进行链延长反应,链延长菌驯化期大于50 d;在乙醇投加量为9~22 g·L−1条件下,MCFA产率为1.9 g·(L·d)−1,主要产物为正己酸。因此,加入驯化富集的链延长菌可大大缩短链延长的启动期,提高MCFA产率,并有利于更长碳链MCFA的合成。
-
目前,关于有机废物链延长的相关综述主要集中于链延长代谢路径[12]、机理[13]、微生物群落[9]、有机废物转化[4]、产物提取[12]、产物利用[9]等方面。本文从有机废物性质、特征入手,对单一有机废物厌氧发酵液及多种有机废物混合发酵液链延长进行了系统分析,提出了有机废物混合发酵液链延长的优势,以期为今后有机废物发酵液链延长研究提供参考。
-
1)污泥。剩余污泥作为污水处理过程的副产品,如果处理不当,会对大气、水体和土壤造成二次污染[36]。有研究表明,剩余污泥的干基热值范围为5 844~19 303 kJ·kg−1[36],能够产生人均150 kWh·a−1的能量[1]。剩余污泥中含有大量的有机物,从糖类、蛋白质等成分上看,剩余污泥与餐厨垃圾等生物质废物较为接近,具有厌氧发酵产酸的资源化前景,同时亦可实现经济效益和产品的多样化。近十几年来,研究者多利用剩余污泥水解酸化生产甲烷或短链有机羧酸,也有部分研究者将污泥厌氧发酵液继续添加到链延长系统中生产己酸,并取得了良好的效果[10]。WU等[1]以乙醇作为电子供体,将含固率16.8 g·L−1的污泥的碱性发酵液作为电子受体,以探究不同电子供体/电子受体对MCFA生产的影响,反应器有效容积为10 L。其研究结果表明:当电子供体/电子受体为1∶1时,己酸质量浓度为2.35 g·L−1,且系统中MCFA占总有机碳的比例达到最高,为72.9%;当电子供体/电子受体为2∶1时,得到的己酸质量浓度最高,为2.39 g·L−1,但系统中MCFA占总有机碳的比例为65.9%。WANG等[11]探究了污泥含固率(2%~8%)对厌氧发酵及后续发酵液链延长产己酸的影响,结果表明,含固率为8%污泥的水解酸化液链延长己酸产量最高,己酸的质量浓度可达到5.04 g·L−1。苑荣雪[37]在体积为200 mL的反应器内以剩余污泥发酵液为底物,进行厌氧发酵链延长实验生产MCFA;结果表明,当底物浓度为600 mmol·L−1时,迟滞期最短,高浓度的乙醇和MCFA均未对链延长微生物产生明显的抑制作用,在醇酸质量比为2∶1的条件下,己酸的质量浓度高达6.20 g·L−1。
2)餐厨垃圾。餐厨垃圾含水率较高、有机质含量丰富,同时富含氮、磷、钾等营养元素,是一种较为合适的厌氧发酵底物[3]。吴云[38]探究了pH对餐厨垃圾厌氧水解酸化的影响,结果表明:当pH为4~5时,发酵类型为乙酸型发酵;当pH为5~6时,发酵类型为丙酸型发酵;当pH为8~9时,发酵类型为丁酸型发酵;当pH在6.5~7.5范围内时,VFA的产量最高,VFA的质量浓度可达46.23 g·L−1。餐厨垃圾还可以在乳酸型发酵过程中生产乳酸,乳酸既可以作为电子供体也可以作为电子受体参与链延长反应,因而可大大提高链延长的反应效率,同时减少乙醇的消耗[2]。王冰[2]探究了不同底物浓度下餐厨垃圾厌氧发酵的乳酸产量,结果表明,当底物质量浓度为30 g·L−1时,乳酸质量分数较高,为505 mg·g−1。WU等[34]发现,当乳酸和乙醇共存时,MCFA产量显著增加。REDDY等[29]将40 mL餐厨垃圾加入120 mL反应器中进行乳酸发酵,得到的乳酸发酵液和乙醇共同投加到链延长系统中,得到己酸质量浓度达到8.10 g·L−1。CONTRERAS-DAVILA等[39]利用餐厨垃圾乳酸发酵液进行了链延长实验,结果表明,随着乳酸的消耗,己酸的质量浓度从1.20 g·L−1提高至5.40 g·L−1。对比上述研究可以发现,当向乳酸发酵液中添加乙醇进行链延长反应时,己酸浓度可得到大幅度提升。
3)果蔬垃圾。果蔬垃圾是在果蔬收获、加工、运输等过程中产生的有机固体废物,具有易于生物降解的特点。果蔬垃圾主要成分为:糖类和半纤维素(75%)、纤维素(9%)和木质素(5%)。果蔬垃圾含水率高、有机质中可溶性物质比例高,因此具有较高的生物可降解性[8]。正是由于果蔬垃圾的这些特点,使厌氧发酵产酸成为果蔬垃圾资源化的有效技术[33]。吴远远[40]研究了pH和有机负荷对果蔬垃圾混合菌群厌氧发酵产物分布的影响,结果表明,在pH为4的条件下,乙醇和乳酸是果蔬垃圾厌氧发酵的主要产物,生成的CO2占气体总量99%以上,可以视为乙醇-乳酸型发酵。郑明月等[41]发现,在果蔬垃圾厌氧发酵过程中,pH对果蔬垃圾发酵类型有显著影响,当pH为4.0时为乙醇型发酵,pH为5.0时为丙酸型发酵,pH为6.0时转变为丁酸型发酵;在最适条件下,发酵产物中乙醇、丙酸和丁酸质量浓度分别为10.5、2.7、8.1 g·L−1。果蔬垃圾厌氧发酵液也可以添加到链延长系统生产己酸等具有更高附加值的产品。YU等[27]利用16.7 g·L−1 (以VS计)的果蔬废物在接种物/底物(质量比)为2∶1的条件下,在有效容积1 L的发酵罐内进行厌氧发酵,厌氧发酵液中乙醇、乙酸、丁酸的质量浓度分别为0.8、5.3、3.6 g·L−1;将厌氧发酵液投加到链延长系统中,同时按照乙醇/酸(质量比)为4∶1的比例继续加入乙醇,在pH为7.5的条件下生产的己酸质量浓度达到14.9 g·L−1。刘春梅[8]提出,合理控制醇酸比和pH可以提高链延长过程的发酵能力,发酵罐容积为1 L,当乙醇和果蔬垃圾发酵液比例(质量比)为4∶1、pH为7.5时,生成的己酸质量浓度最高达到14.9 g·L−1。同时,刘春梅使用离子交换树脂提取己酸,可有效降低己酸积累对链延长系统微生物的毒性。通过己酸提取可使反应过程中的己酸质量浓度提高至17.31 g·L−1,最高己酸产率为2.02 g·(L·d)−1。
4)木质纤维素类有机废物。农业(玉米秸秆、稻草等)、林业(林木、树枝、叶片等)、能源作物(柳条草、黄杨等)、市政绿化(草、灌木等)等积累了大量的木质纤维素类废物[40]。这些有机废物中含有大量的有机物,主要为纤维素、半纤维素、木质素,其所占比例分别为30%~45%、25%~40%、10%~20%[42]。WANG等[43]在100 mL有效容积的血清瓶中用瘤胃液对草坪草进行发酵,以提高草坪草中有机物的水解酸化;结果表明,当草坪草含固率为5%时,发酵液中VFA的质量浓度为10.2 g·L−1。近年来,也有研究者将木质纤维素类废物厌氧发酵液作为电子受体进行链延长产己酸实验,并取得了较好的效果。ZHANG等[44]研究了麦秸、草坪草、木屑3种木质纤维素类废物厌氧发酵液在100 mL血清瓶中链延长生产己酸的效果,己酸产量从高到低的顺序为麦秸>草坪草>木屑;含固率为40 g·L−1的麦秸发酵液经过链延长生成的己酸质量浓度最高达到8.61 g·L−1,不仅麦秸表面而且麦秸内部多孔结构均可为微生物生长繁殖提供巨大的比表面积,并且在麦秸上形成具有较高活性和稳定性的生物膜,对氨抑制具有极高的耐受性,因此,以麦秸作为载体的生物膜在己酸生产过程中可重复利用。WEIMER等[45]利用瘤胃液对苜蓿茎和柳枝稷等木质纤维素类有机废物进行厌氧发酵(发酵液体积为160 mL),产生了少量的戊酸和己酸;之后,在系统中加入克氏梭菌,戊酸和己酸的产量显著增加;再加入乙醇进行专门的链延长,得到的丁酸、戊酸、己酸的质量浓度分别为2.3、1.6、6.1 g·L−1。KENEALY等[46]将瘤胃纤维分解菌和克氏梭菌结合,投加到60 mL血清瓶中,使其在纤维素和乙醇共存的体系中共同生长,利用木质纤维素类有机废物同时进行厌氧发酵和链延长,最终获得的己酸质量浓度为4.6 g·L−1。KHOR等[47]先将草在浸出床反应器中发酵生成乳酸,再在100 mL的二级发酵罐中采用电辅助的方式进行链延长生产己酸,己酸质量浓度在pH=5.5时可达到10.92 g·L−1。将己酸产物以阴离子的形式提取,浓缩并通过膜电解转化,可得到纯度为70%的己酸溶液,获得了迄今为止木质纤维素类废物为原料的最高己酸生产速率,约为0.99 g·(L·d)−1。
无论是污泥、餐厨垃圾、果蔬垃圾还是木质纤维素类有机废物,其厌氧发酵液链延长生产MCFA,均能有效利用生物质。由于污泥、餐厨垃圾和果蔬垃圾等有机物丰富、含糖量较高,其厌氧发酵液链延长所产己酸较多,目前对其研究亦较多。而木质纤维素类生物质与前面几种有机废物相比,含有更多的有机大分子聚合物,生物降解性能较差,其厌氧发酵和链延长产己酸的效率也较低。因此,将这些底物进行混合发酵及链延长,可实现基质的互相补充,对提高己酸产量和产率均具有重要意义。
-
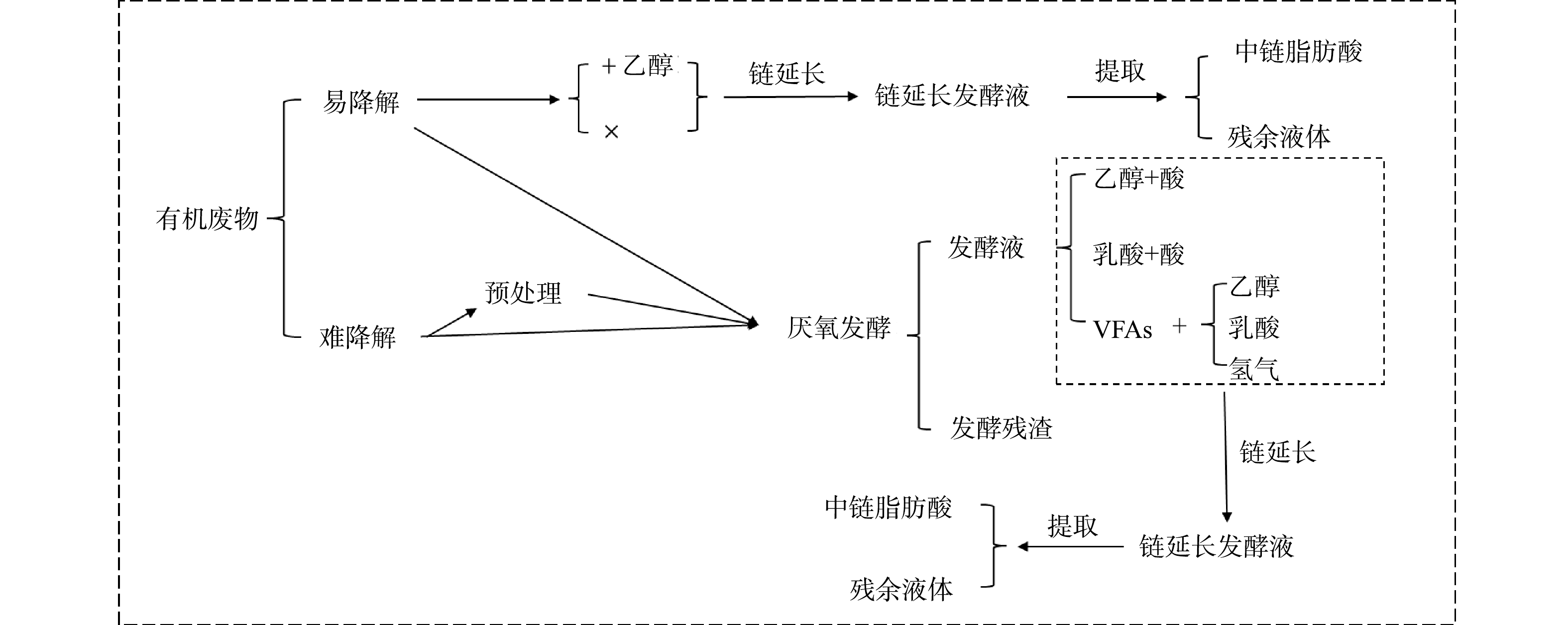
将2种或多种有机底物一起进行厌氧发酵的方式称为共发酵或混合发酵[48]。2种或2种以上有机底物共发酵可以实现碳源和氮源的互相补充。例如,于佳动[42]从C/N比的角度将木质纤维素类废物分为高C/N比底物(如秸秆)和低C/N比底物(如禽畜粪便等)。C/N比过高可使体系缓冲能力降低,反应过程不稳定,不利于微生物的生长;而C/N比过低容易引起氨抑制,也会造成厌氧发酵体系不稳定。因此,将高C/N比和低C/N比的原料按比例混合,可将C/N比控制在适宜范围,从而获得更好的发酵效果。李月寒[49]在1 L反应器中通过超声破解促进污泥和餐厨垃圾的共发酵,经过产酸条件优化得到的VFA质量浓度为29.23~30.78 g·L−1,而超声预处理剩余污泥的产酸质量浓度仅为6.944 g·L−1。有研究表明,污泥与餐厨垃圾[50]、污泥与秸秆[3]、餐厨垃圾与秸秆[3]、混合的城市有机固体废物[7]共发酵均可产生比其单一有机底物水解酸化更高浓度的VFAs(图3)。
餐厨垃圾自身碳水化合物含量高、C/N比高,所含微生物种类可能较少[2];而剩余污泥C/N比低、并且含有丰富的微生物[9]。两者共发酵会具有协同作用:一方面可以利用餐厨垃圾中丰富的有机物和剩余污泥中丰富的氮源实现物料互补、营养平衡,得到更高的VFA产量;另一方面,可以利用污泥中丰富的微生物群落提高共发酵系统的运行效率和稳定性[50]。王冰[2]将餐厨垃圾和剩余污泥在1 L反应器中进行厌氧共发酵,发酵液中富含乳酸,将发酵液添加至链延长系统中作为电子供体,系统中丁酸和戊酸的浓度增加,但没有生成己酸;当继续投加乙醇使其与乳酸形成联合电子供体时,生成的己酸浓度大幅度升高,经过10 d的链延长反应,己酸的累积质量浓度达到8.3 g·L−1,比剩余污泥水解酸化液和餐厨垃圾水解酸化液链延长的最高己酸质量浓度分别高2.1和0.3 g·L−1[1, 8]。
城市有机固体废物包含绿色有机废物(如个人住宅的花园垃圾(较难降解))、食物垃圾(如一部分厨房垃圾和蔬菜垃圾(易降解)),以及一些难降解的塑料和其他剩余垃圾(纸巾、烟头、布料等)[9]。城市固体废物中的有机组分可通过共发酵和链延长技术转化为MCFA。GROOTSCHOLTEN等[20]在5 L单相厌氧发酵体系和两相厌氧发酵体系中分别研究了以城市有机固体废物为原料的发酵产酸和链延长产己酸效果,己酸产率分别为0.7 g·(L·d)−1和1.9 g·(L·d)−1,得到的己酸质量浓度分别为2.7 g·L−1和12.6 g·L−1。可以看出,不论己酸产率还是己酸浓度,两相厌氧发酵-链延长效果均优于单相厌氧发酵体系。
-
以有机废物厌氧发酵液作为底物进行链延长生产MCFA是实现有机废物资源化的良好途径。目前,单一有机废物厌氧发酵液作为底物进行链延长生产己酸已经具备了一定的研究基础并形成了理论体系,但有关多种有机废物共发酵进行链延长的研究还处于起步阶段。现阶段生物合成己酸的研究中通常使用酵母浸粉提供氮源,成本较高。而在多有机底物共发酵链延长过程中,可以实现氮源、碳源的互相补充,提高己酸产量,并降低生产成本。未来的相关研究应该更加注重以下几个方面。
1)相比于以单一有机废物发酵液为底物进行链延长反应,以多种有机物混合发酵时,其发酵液组分更为复杂,微生物亦更加多样。因此,多种有机废物混合发酵的特征,及其以该发酵液作为底物进行链延长的条件优化、微生物群落及其反应机制等,需要进一步探究。
2)在微生物链延长过程中,乙醇是良好的电子供体,直接添加化学乙醇是常用方法,而通过有机废物厌氧发酵生产生物质乙醇用于链延长反应对于推进废物资源化也非常有必要。如何协调厌氧发酵产乙醇、产VFA和后续微生物链延长产MCFA之间的关系,是实现高效经济MCFA生产的重要基础,需要进一步深入研究。
3)目前对于链延长的研究多采用序批式反应器,无法在反应过程中及时监测和调控pH。此外,链延长生成的MCFA也可能反过来抑制链延长过程。因此,将连续流厌氧发酵反应器应用于链延长需深入开展研究。
4)将有机废物厌氧发酵-链延长产MCFA进行中试和生产性实验,是实现该技术推广应用的重要环节,仍需进一步开展研究。同时,有机废物厌氧发酵-链延长产MCFA的经济性评价和全生命周期评价也非常重要。
5)利用有机废物发酵液进行链延长反应,丰富了链延长生产己酸的体系和己酸生产效率,但链延长体系中的高浓度己酸也对链延长功能的微生物有一定抑制作用。虽然己酸比VFA有更好的分离性能,但从发酵液中分离和纯化己酸仍然是高附加值产品生产的一个关键,需要进一步深入开展研究。
有机废物厌氧发酵液链延长合成中链脂肪酸研究进展
Study progress on chain elongation of anaerobic fermentation liquid from organic wastes for medium-chain fatty acid synthesis
-
摘要: 有机废物产量巨大,具备较高的资源化利用潜力。以有机废物厌氧发酵液为底物,通过链延长工艺生产中链脂肪酸,可有效提升产物的经济价值,因而是一种极具潜力的有机废物资源化方法。梳理了有机废物厌氧发酵液链延长的机理以及影响因素,分析了不同有机废物在厌氧发酵链延长中的作用,并提出了今后研究的方向。学术界已获得的研究结果表明,以混合有机废物作为底物进行厌氧发酵,可以实现氮源和碳源的相互补充,其获得的发酵液中挥发性脂肪酸浓度更高,这对后续的链延长产中链脂肪酸具有良好的促进作用。Abstract: The production of organic wastes is huge, presenting a high potential of resource utilization. The production of medium chain fatty acid (MCFA) by chain elongation process using organic wastes anaerobic fermentation liquid as substrate can effectively enhance the economic value of the products, thus is a highly promising method for resource utilization of organic waste. The mechanisms and influencing factors of chain elongation from organic waste fermentation liquids were combed, the role of different organic waste in anaerobic fermentation and further chain elongation was analyzed, the directions for future research were also proposed. The results of studies that have been obtained in academia showed that mixed organic wastes as substrate of anaerobic fermentation can achieve mutual complementation of nitrogen and carbon source, and the higher volatile fatty acid concentration was achieved in the fermentation liquid, which can promote the subsequent chain elongation and MCFA production.
-

-
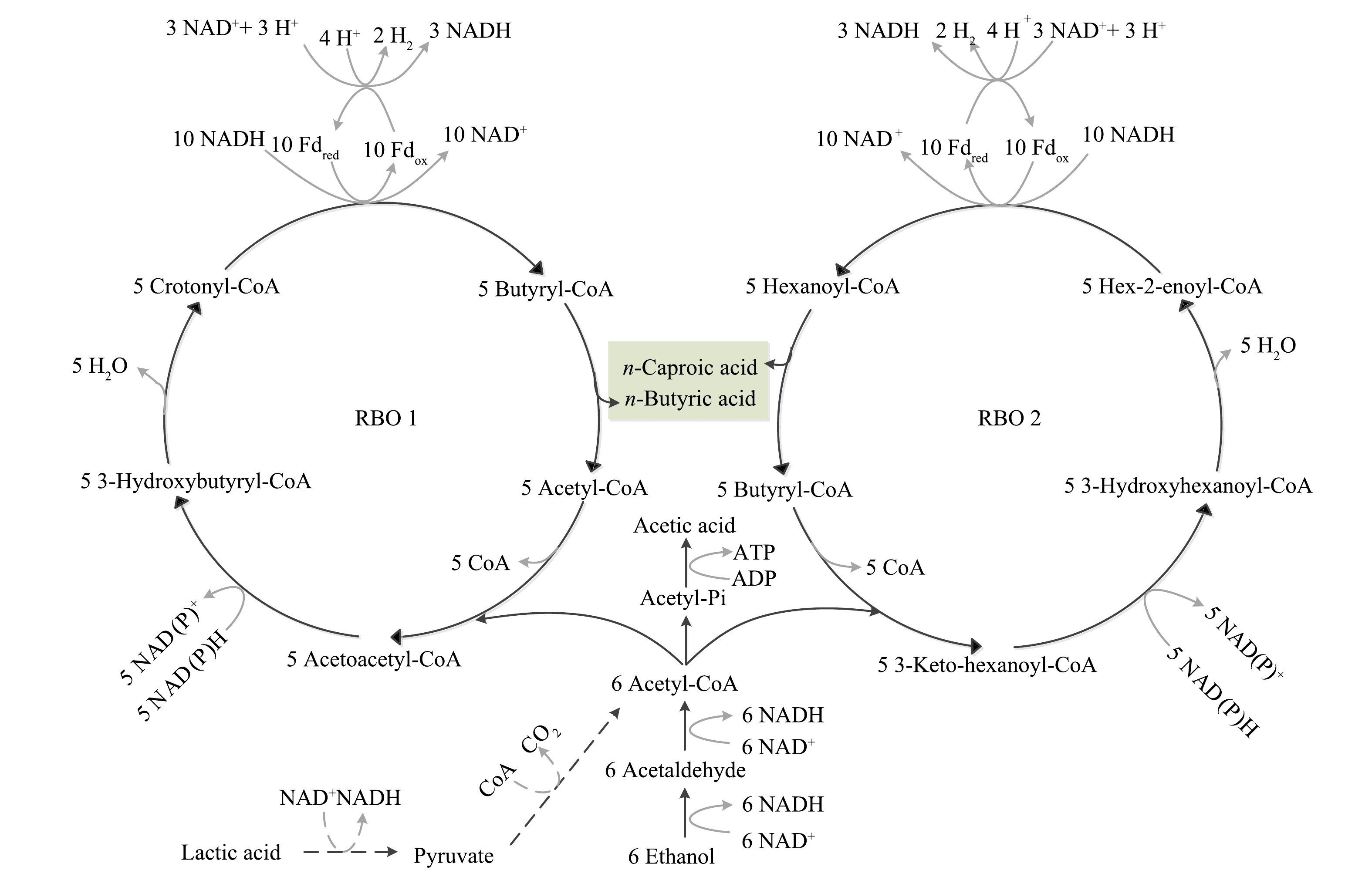
图 1 克氏梭菌作用下羧酸链延长代谢路径[9]
Figure 1. Metabolic pathways of carboxylic acid chain elongation by Clostridium kluyveri
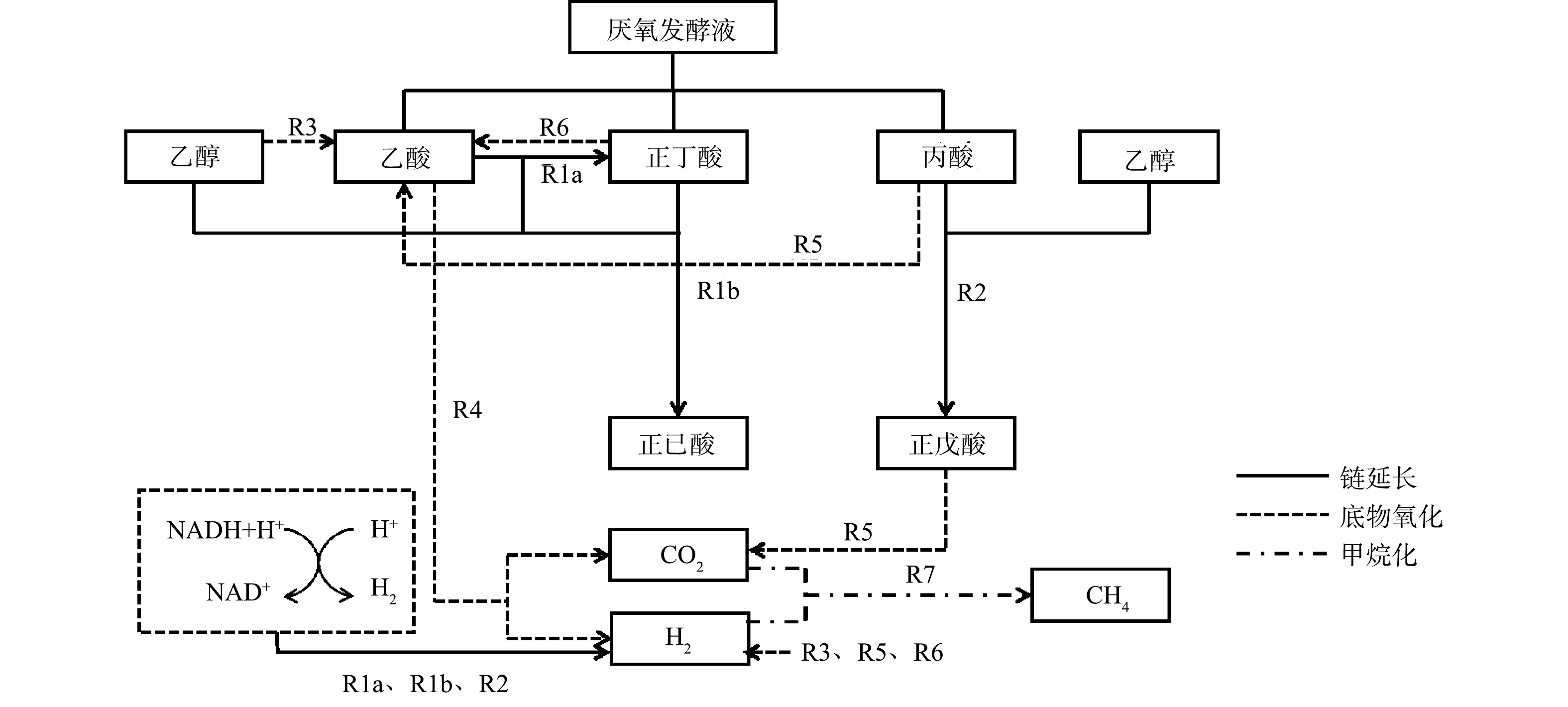
图 2 基于克氏梭菌的有机废物厌氧发酵液链延长的合成代谢和分解代谢路径[11]
Figure 2. Anabolic and catabolic pathways of chain elongation of organic waste anaerobic fermentation liquid with Clostridium kluyveri
表 1 有机废物厌氧发酵液链延长和氧化的主要代谢反应
Table 1. Main metabolic reactions of chain elongation and oxidation of organic waste anaerobic fermentation liquid
编号 路径 反应式 R1a 乙酸→正丁酸 $\text{6ethanol+4}{\text{acetate} }^{\text{-} }\text{→}\text{5}{{n}\text{–butyrate} }^{\text{-} }\text{+}{\text{H} }^{\text{+} }\text{+2}{\text{H} }_{\text{2} }\text{+4}{\text{H} }_{\text{2} }\text{O}$ R1b 正丁酸→正己酸 $\text{6ethanol+5}{{n}\text{–butyrate} }^{\text{-} }\text{→}\text{5}{{n}\text{–caproate} }^{\text{-} }\text{+}{\text{acetate} }^{\text{-} }\text{+}{\text{H} }^{\text{+} }\text{+2}{\text{H} }_{\text{2} }\text{+4}{\text{H} }_{\text{2} }\text{O}$ R2 丙酸→正戊酸 $\text{6ethanol+5}{\text{propionate} }^{\text{-} }\text{→}\text{5}{{n}\text{–valerate} }^{\text{-} }\text{+}{\text{acetate} }^{\text{-} }\text{+}{\text{H} }^{\text{+} }\text{+2}{\text{H} }_{\text{2} }\text{+4}{\text{H} }_{\text{2} }\text{O}$ R3 乙醇氧化 $ \text{ethanol+}{\text{H}}_{\text{2}}\text{O}\text{→}{\text{acetate}}^{\text{-}}\text{+}{\text{H}}^{\text{+}}\text{+2}{\text{H}}_{\text{2}} $ R4 乙酸氧化 $ {\text{acetate}}^{\text{-}}\text{+}{\text{H}}^{\text{+}}\text{+2}{\text{H}}_{\text{2}}\text{O}\text{→}\text{2}{\text{CO}}_{\text{2}}\text{+4}{\text{H}}_{\text{2}} $ R5 丙酸氧化 $ {\text{propionate}}^{\text{-}}\text{+2}{\text{H}}_{\text{2}}\text{O}\text{→}{\text{acetate}}^{\text{-}}\text{+C}{\text{O}}_{\text{2}}\text{+3}{\text{H}}_{\text{2}} $ R6 正丁酸氧化 ${{n}\text{–butyrate} }^{\text{-} }\text{+2}{\text{H} }_{\text{2} }\text{O}\text{→}\text{2}{\text{acetate} }^{\text{-} }\text{+}{\text{H} }^{\text{+} }\text{+2}{\text{H} }_{\text{2} }$ R7 甲烷化 $ {\text{CO}}_{\text{2}}\text{+4}{\text{H}}_{\text{2}}\text{→}{\text{CH}}_{\text{4}}\text{+2}{\text{H}}_{\text{2}}\text{O} $  下载: 导出CSV
下载: 导出CSV
表 2 利用开放的链延长系统生产己酸的效果
Table 2. Effectiveness of caproic acid production in open chain elongation system
底物 反应器类型 pH 己酸质量浓度/ (g·L−1) 己酸产率/(g·(L·d)−1) 文献 乙酸、乙醇 序批式反应器 7.0 8.2 0.48 [23] 乙酸、乙醇 序批式反应器 6.75~7.0 4.82 — [16] 剩余污泥水解酸化液、乙醇 序批式反应器 6.75~6.8 5.04 — [11] 办公室碎纸、鸡粪 序批式反应器 — 10 — [28] 餐厨垃圾、乙醇 序批式反应器 7.0 8.1 — [29] 乙酸、乙醇 上流式反应器 6.5~ 7.0 11.1 15.7 [19] 乙酸、乙醇 上流式反应器 5.5~7.0 21.1 — [24] 乙酸、乙醇 上流式反应器 6.5~7.2 12.0 57.4 [30] 活性污泥碱性发酵液、乙醇 上流式反应器 7.1 2.39 — [1] 城市固体废物、乙醇 两相补料间歇式厌氧反应器 6.5~7.0 12.6 26 [7] 玉米啤酒废水 半批式反应器 5.5 — 3.4 [31]
下载: 导出CSV
-
[1] WU S L, SUN J, CHEN X M, et al. Unveiling the mechanisms of medium-chain fatty acid production from waste activated sludge alkaline fermentation liquor through physiological, thermodynamic and metagenomic investigations[J]. Water Research, 2019, 169: 115218. [2] 王冰. 餐厨垃圾厌氧发酵产中链脂肪酸的研究[D]. 哈尔滨: 哈尔滨工业大学, 2019 [3] 方卉, 赵剑斐, 彭道平, 等. 秸秆混合厌氧发酵研究进展[J]. 四川环境, 2019, 38(3): 187-192. [4] 朱文彬. 有机废物厌氧发酵生物合成己酸研究进展[J]. 环境工程, 2020, 38(1): 128-134. [5] AGLER M T, SPIRITO C M, USACK J G, et al. Chain elongation with reactor microbiomes: Upgrading dilute ethanol to medium-chain carboxylates[J]. Energy & Environmental Science, 2012, 5(8): 8189-8192. [6] ANGENENT L T, RICHTER H, BUCKEL W, et al. Chain elongation with reactor microbiomes: Open-culture biotechnology to produce biochemicals[J]. Environmental Science & Technology, 2016, 50(6): 2796-2810. [7] GROOTSCHOLTEN T I M, STRIK D P B T B, STEINBUSCH K J J, et al. Two-stage medium chain fatty acid (MCFA) production from municipal solid waste and ethanol[J]. Applied Energy, 2014, 116: 223-229. doi: 10.1016/j.apenergy.2013.11.061 [8] 刘春梅. 两相法厌氧发酵产己酸及其微生物学研究[D]. 无锡: 江南大学, 2018 [9] CAVALCANTE W D A, LEITAO R C, GEHRING T A, et al. Anaerobic fermentation for n-caproic acid production: A review[J]. Process Biochemistry, 2017, 54: 106-119. doi: 10.1016/j.procbio.2016.12.024 [10] 陈哲柯. 基于厌氧发酵的剩余污泥产中链脂肪酸研究[D]. 长沙: 湖南大学, 2018 [11] WANG Q, ZHANG P, BAO S, et al. Chain elongation performances with anaerobic fermentation liquid from sewage sludge with high total solid as electron acceptor[J]. Bioresource Technology, 2020, 306: 123188. doi: 10.1016/j.biortech.2020.123188 [12] WU Q, JIANG Y, CHEN Y, et al. Opportunities and challenges in microbial medium chain fatty acids production from waste biomass[J]. Bioresource Technology, 2021, 340: 125633. doi: 10.1016/j.biortech.2021.125633 [13] WU Q, BAO X, GUO W, et al. Medium chain carboxylic acids production from waste biomass: Current advances and perspectives[J]. Biotechnology Advance, 2019, 37(5): 599-615. doi: 10.1016/j.biotechadv.2019.03.003 [14] LIU Y, LU F, SHAO L, et al. Alcohol-to-acid ratio and substrate concentration affect product structure in chain elongation reactions initiated by unacclimatized inoculum[J]. Bioresource Technology, 2016, 218: 1140-1150. doi: 10.1016/j.biortech.2016.07.067 [15] YIN Y, ZHANG Y, KARAKASHEV D B, et al. Biological caproate production by Clostridium kluyveri from ethanol and acetate as carbon sources[J]. Bioresource Technology, 2017, 241: 638-644. doi: 10.1016/j.biortech.2017.05.184 [16] BAO S, WANG Q, ZHANG P, et al. Effect of acid/ethanol ratio on medium chain carboxylate production with different VFAs as the electron acceptor: Insight into carbon Balance and microbial community[J]. Energies, 2019, 12(19): 3720. doi: 10.3390/en12193720 [17] 包率. 污泥水解酸化液链延长混合发酵产中链脂肪酸研究[D]. 北京: 北京林业大学, 2019 [18] AGLER M T, SPIRITO C M, USACK J G, et al. Development of a highly specific and productive process for n-caproic acid production: Applying lessons from methanogenic microbiomes[J]. Water Science and Technology, 2014, 69(1): 62-68. doi: 10.2166/wst.2013.549 [19] GROOTSCHOLTEN T I, STEINBUSCH K J, HAMELERS H V, et al. Chain elongation of acetate and ethanol in an upflow anaerobic filter for high rate MCFA production[J]. Bioresource Technology, 2013, 135: 440-445. doi: 10.1016/j.biortech.2012.10.165 [20] GROOTSCHOLTEN T I M, KINSKY D B F, HAMELERS H V M, et al. Promoting chain elongation in mixed culture acidification reactors by addition of ethanol[J]. Biomass and Bioenergy, 2013, 48: 10-16. doi: 10.1016/j.biombioe.2012.11.019 [21] 卓英莲. 污水厂剩余污泥水解酸化产挥发性脂肪酸的试验研究[D]. 广州: 华南理工大学, 2010 [22] WEIMER P J, STEVENSON D M. Isolation, characterization, and quantification of Clostridium kluyveri from the bovine rumen[J]. Applied Microbiology and Biotechnology, 2011, 94(2): 461-466. [23] STEINBUSCH K J J, HAMELERS H V M, PLUGGE C M, et al. Biological formation of caproate and caprylate from acetate: Fuel and chemical production from low grade biomass[J]. Energy& Environmental Science, 2011, 4(1): 216-224. [24] LIU Y, HE P, SHAO L, et al. Significant enhancement by biochar of caproate production via chain elongation[J]. Water Research, 2017, 119: 150-159. doi: 10.1016/j.watres.2017.04.050 [25] VASUDEVAN D, RICHTER H, ANGENENT L T. Upgrading dilute ethanol from syngas fermentation to n-caproate with reactor microbiomes[J]. Bioresource Technology, 2014, 151: 378-382. doi: 10.1016/j.biortech.2013.09.105 [26] BOLAJI I O, DIONISI D. Acidogenic fermentation of vegetable and salad waste for chemicals production: Effect of pH buffer and retention time[J]. Journal of Environmental Chemical Engineering, 2017, 5(6): 5933-5943. doi: 10.1016/j.jece.2017.11.001 [27] YU J, HUANG Z, WU P, et al. Performance and microbial characterization of two-stage caproate fermentation from fruit and vegetable waste via anaerobic microbial consortia[J]. Bioresource Technology, 2019, 284: 398-405. doi: 10.1016/j.biortech.2019.03.124 [28] LONKAR S, FU Z, HOLTZAPPLE M. Optimum alcohol concentration for chain elongation in mixed-culture fermentation of cellulosic substrate[J]. Biotechnology and Bioengineering, 2016, 113(12): 2597-2604. doi: 10.1002/bit.26024 [29] REDDY M V, HAYASHI S, CHOI D, et al. Short chain and medium chain fatty acids production using food waste under non-augmented and bio-augmented conditions[J]. Journal of Cleaner Production, 2018, 176: 645-653. doi: 10.1016/j.jclepro.2017.12.166 [30] GROOTSCHOLTEN T I, STEINBUSCH K J, HAMELERS H V, et al. Improving medium chain fatty acid productivity using chain elongation by reducing the hydraulic retention time in an upflow anaerobic filter[J]. Bioresource Technology, 2013, 136: 735-738. doi: 10.1016/j.biortech.2013.02.114 [31] GE S, USACK J G, SPIRITO C M, et al. Long-term n-caproic acid production from yeast-fermentation beer in an anaerobic bioreactor with continuous product extraction[J]. Environmental Science & Technology, 2015, 49(13): 8012-8021. [32] 石耀威. 厌氧微生物转化木薯酒糟制取中链脂肪酸工艺及机理研究[D]. 北京: 北京化工大学, 2020 [33] KUCEK L A, NGUYEN M, ANGENENT L T. Conversion of L-lactate into n-caproate by a continuously fed reactor microbiome[J]. Water Research, 2016, 93: 163-171. doi: 10.1016/j.watres.2016.02.018 [34] WU Q, GUO W, YOU S, et al. Concentrating lactate-carbon flow on medium chain carboxylic acids production by hydrogen supply[J]. Bioresource Technology, 2019, 291: 121573. [35] SPIRITO C M, RICHTER H, RABAEY K, et al. Chain elongation in anaerobic reactor microbiomes to recover resources from waste[J]. Current Opinion in Biotechnology, 2014, 27: 115-122. doi: 10.1016/j.copbio.2014.01.003 [36] 王园园, 张光明, 张盼月, 等. 污泥厌氧发酵制氢研究进展[J]. 水资源保护, 2016, 32(4): 109-116. doi: 10.3880/j.issn.1004-6933.2016.04.018 [37] 苑荣雪. 以污泥发酵液为底物产中链脂肪酸可行性研究[J]. 环境科学与技术, 2019, 42(11): 141-146. [38] 吴云. 餐厨垃圾厌氧消化影响因素及动力学研究[D]. 重庆: 重庆大学, 2009 [39] CONTRERAS-DAVILA C A, CARRION V J, VONK V R, et al. Consecutive lactate formation and chain elongation to reduce exogenous chemicals input in repeated-batch food waste fermentation[J]. Water Research, 2019, 169: 115215. [40] 吴远远. 基于厌氧产酸发酵的新型果蔬垃圾厌氧处理工艺研究[D]. 北京: 清华大学, 2016. [41] 郑明月, 郑明霞, 王凯军, 等. 温度、pH和负荷对果蔬垃圾厌氧酸化途径的影响[J]. 可再生能源, 2012, 30(4): 75-79. [42] 于佳动. 纤维质农业废弃物微好氧酸化机理及高含固率两相厌氧发酵工艺研究[D]. 北京: 中国农业大学, 2017. [43] WANG S, ZHANG G, ZHANG P, et al. Rumen fluid fermentation for enhancement of hydrolysis and acidification of grass clipping[J]. Journal of Environmental Management, 2018, 220: 142-148. [44] ZHANG C, YANG L, TSAPEKOS P, et al. Immobilization of Clostridium kluyveri on wheat straw to alleviate ammonia inhibition during chain elongation for n-caproate production[J]. Environment International, 2019, 127: 134-141. doi: 10.1016/j.envint.2019.03.032 [45] WEIMER P J, NERDAHL M, BRANDL D J. Production of medium-chain volatile fatty acids by mixed ruminal microorganisms is enhanced by ethanol in co-culture with Clostridium kluyveri[J]. Bioresource Technology, 2015, 175: 97-101. doi: 10.1016/j.biortech.2014.10.054 [46] KENEALY W R, CAO Y, WEIMER P J. Production of caproic acid by cocultures of ruminal cellulolytic bacteria and Clostridium kluyveri grown on cellulose and ethanol[J]. Applied Microbiology Biotechnology, 1995, 44: 507-513. doi: 10.1007/BF00169952 [47] KHOR W C, ANDERSEN S, VERVAEREN H, et al. Electricity-assisted production of caproic acid from grass[J]. Biotechnology for Biofuels, 2017, 10: 180. doi: 10.1186/s13068-017-0863-4 [48] 边道林, 孙磊, 王爽, 等. 多底物共发酵产酸特性研究[J]. 黑龙江农业科学, 2014(9): 28-31. doi: 10.3969/j.issn.1002-2767.2014.09.007 [49] 李月寒. 超声波预处理污泥和厨余共发酵产短链脂肪酸的研究[D]. 苏州: 苏州科技大学, 2016 [50] 郑舍予. 剩余污泥联合餐厨垃圾高温共发酵产酸研究[D]. 上海: 华东理工大学, 2019 -
 点击查看大图
点击查看大图
计量
- 文章访问数: 6164
- HTML全文浏览数: 6164
- PDF下载数: 333
- 施引文献: 0
