



 下载:
下载:













-
榨菜加工过程中产生的废水含高浓度有机物(以COD计为300~2 000 mg·L−1)、高盐度(以NaCl计为15~25 g·L−1)与高氨氮(200~300 mg·L−1)[1-2]。此类废水以及其他高盐废水需高效处理后才能排放,否则将对土壤和水体环境造成极大的威胁[3]。厌氧和好氧技术通常被联合用于高盐废水治理[4-6]。然而,综合处理过程不但消耗了大量能量、易造成二次污染,而且常常不能同时达到氮和碳的排放标准[7]。值得注意的是,生物电化学系统(biological electrochemical system,BES)是一种可持续且具有成本效益的技术,已证明具有良好脱氮能力[8-10]。BES在处理C/N为0的含氮废水时,依然有着较高的硝酸盐去除率[11]。
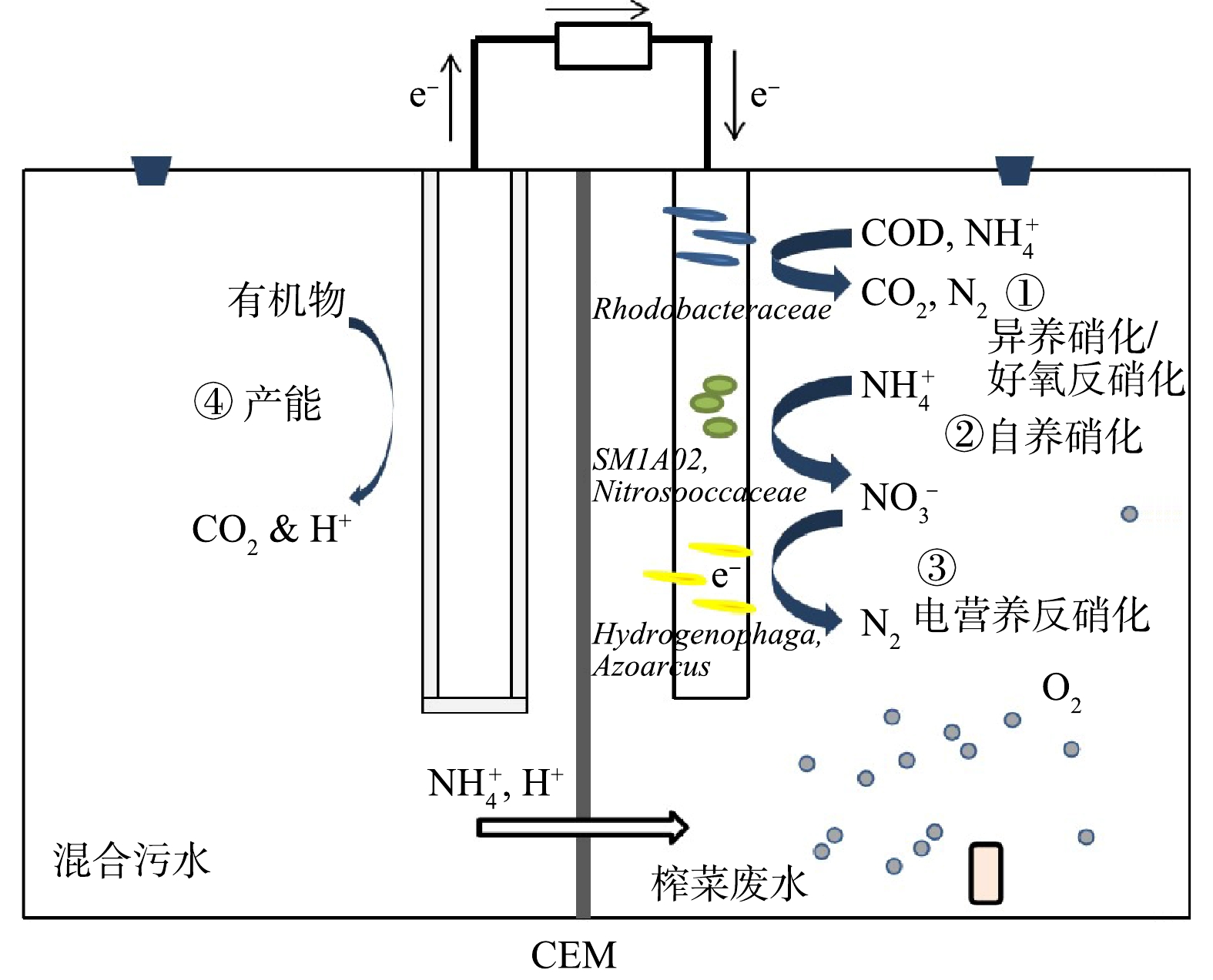
就功能而言,混合生物阴极 MFC可实现BES中不同形式氮的转化与去除[11]。用厌氧泥与好氧泥依次混合接种MFC阴极的方式可实现良好的TN去除率(89.8%~97.6%)[12]。最近有研究[13]表明,混合生物阴极 MFC在处理榨菜废水时实现了完全脱氮,并提出脱氮机理包括盛宴阶段、饥荒阶段和稳定阶段共3个阶段的理论。
在盛宴阶段、饥荒阶段与稳定阶段发生的主反应分别为异养硝化/好氧反硝化反应、自养硝化与电营养反硝化[13]。异养硝化/好氧反硝化菌可利用基底中的有机物作为电子供体,将不同形态的氮转化成N2[14]。异养硝化菌在碳源充足的条件下,将含氮的化合物氧化成
${\rm{NO}}_2^ - $ 、${\rm{NO}}_3^ - $ ,且大部分能在曝气的条件下将硝态氮还原成氮气,实现好氧反硝化[15]。相较于传统脱氮工艺,异养硝化/好氧反硝化可在同一个反应器中进行,且具有更高的氨氮去除效率[16]。异养硝化反应所产生的氨氮可作为异养硝化/好氧反硝化菌的反应物,从而实现同步硝化和反硝化效果[17]。目前,被报道出的异养硝化/好氧反硝化菌(属)有Paracoccus、Thauera、unclassified_f__Rhodobacteraceae、Flavobacterium、Arcobacter、Halomonas[18-19]。电营养反硝化是一种有潜力的脱氮新技术[13]。电营养反硝化一般机理为:阴极生物膜上存在一类电活性自养脱氮微生物,可直接利用电子,将阴极底物中的
${\rm{NO}}_3^ - $ 与${\rm{NO}}_2^ - $ 还原成N2。在生物电极脱氮过程中,电营养反硝化可减少传统反硝化反应对有机物的依赖[20]。目前已报道参与电营养反硝化的潜在菌(属)有Thauera、Acholeplasm、tappia indica、Xanthobacter、Azoarcus、Pseudomonas stutzer[11,15,21]。混合生物阴极 MFC稳定阶段周期较长,主要原因为稳定阶段的电营养反硝化速率缓慢[13]。因此,探究电营养反硝化速率的影响因素,已成为混合膜 MFC走向实践应用所面临的一个关键。目前,关于生物电极脱氮电子传递机制的研究尚少。根据已有的研究,推测的传递方式可能为直接接触方式、电子中介体方式。电营养反硝化菌可以分泌电子介体,且外源电子介体可有效提高生物电极脱氮的效率[21]。非膜结合细胞色素蛋白、Rnf复合体、红素氧还蛋白、氢化酶与甲酸脱氢酶可能参与生物电极脱氮中电子的直接传递[22]。电子传递链的两端分别为阴极电子和硝酸盐,因此,电流强度对双室混合膜 MFC 的电营养反硝化具有直接影响,但关于这方面的研究目前鲜有报道。
为优化混合生物阴极MFC处理高盐榨菜废水时的脱氮效果,本研究通过改变外电阻,设置了4组不同峰值电流强度(S1、S2、S3、S4分别为(0.24±0.03)、(0.37±0.03)、(0.55±0.11)、(0.5±0.2) mA)的实验,探讨了不同电流强度对高盐双室混合膜 MFC脱氮的影响,并分析了对应的产电特性和微生物群落,优化了反应器运行的最佳工况条件,为后续双室混合膜 MFC处理高盐废水研究提供思路与解决方法。
全文HTML




-
阴极的榨菜废水源自重庆市某榨菜废水处理厂,为初沉池出水。阳极液采用调制后的生活污水。实验使用乙酸钠、NH4Cl、NaCl将COD、
${\rm{NH}}_4^ + $ 、盐度分别调至表1所示水质,原始废水中不含${\rm{NO}}_3^ - $ 和${\rm{NO}}_2^ - $ 。调好的实验用水置于4 ℃的冰箱内保存。 -
实验装置如图1所示,装置采用双室构型,阴阳极室的有效容积均为252 mL(长宽高为6 cm×6 cm×7 cm)。阴阳极室被阳离子交换膜隔开,阳离子交换膜的有效膜面积为42 cm2 (长宽为6 cm×7 cm)。阴阳两电极材料均为碳毡,通过钛丝与铜导线相连,装置启动连接外电阻为1 000 Ω,并连接电压采集器。
本研究设置4组不同外接电阻(1 000、500、100、20 Ω)的实验(S1、S2、S3、S4),通过改变外电阻实现闭合电路中不同的电流强度。在曝气阶段,S1、S2、S3、S4双室混合膜 MFC的峰值电流分别为(0.33±0.01)、(0.44±0.03)、(1.39±0.19)、(0.85±0.2) mA;在停曝时期,S1、S2、S3、S4双室混合膜 MFC的峰值电流分别为(0.24±0.03)、(0.37±0.03)、(0.55±0.11)、(0.5±0.2) mA。实验的顺序先后为S1、S2、S3、S4。当总氮去除完毕时,1组实验结束,更换新鲜的阴极液与阳极液,并进行下一组实验。具体接种方式为:先在阳极接种10 mL来自榨菜废水处理厂厌氧池的厌氧污泥,使阳极的产电菌尽快富集并产电,阴极接种10 mL来自榨菜废水处理厂好氧池的好氧污泥,使阴极具有良好的硝化能力[13];待阳极电势高至−400~−500 mV并保持稳定后,再往阴极接种10 mL来自榨菜废水处理厂厌氧池的厌氧污泥,以加强其阴极的反硝化能力[13]。在污泥接种完毕后,待连续2个周期获得稳定的电压和TN的完全去除后,开始正式采集数据。
原理图如图2所示,阳极与阴极均以高盐废水为底物,
${\rm{NH}}_4^ + $ 与H+通过阳离子交换膜在阳极室和阴极室进行交换。阳极中的产电菌消耗高盐废水中的耗氧有机物(以COD计)而产生电流。阴极室中有曝气头,盛宴期氧气与碳源充足,异养硝化/好氧反硝化菌将阴极液中的${\rm{NH}}_4^ + $ -N转化为N2,同时含碳有机物被微生物的生命活动所利用并转化为CO2[18];在饥荒期,碳源不足,自养硝化细菌将残余的${\rm{NH}}_4^ + $ -N转化为硝态氮,且最终转化为${\rm{NO}}_3^ - $ ,过程伴随着一定的内碳源反硝化;稳定期处于无曝气状态,阴极上的电营养反硝化菌直接利用阳极有机物降解产生的电子进行${\rm{NO}}_3^ - $ 的还原,${\rm{NO}}_3^ - $ 被还原成N2,从而达到阴极榨菜废水中总氮的完全去除[13]。阴极前期硝化反应所需要的氧气由气泵提供,曝气量通过手动转动转子流量计阀门调节。曝气速率根据杨茜等[23]的研究,控制在100 mL·min−1,阴极曝气时期溶解氧浓度维持在3~4 mg·L−1。当阴极检测出的氨氮值为0后,停止曝气。本研究中S1、S2、S3、S4停曝时间点分别为第11、9、7、13 d。通过持续给阳极补充乙酸钠试剂以维持电压的恒定,保证充足的电子供给。每次补加,使混合后阳极液的COD大致维持在2 000 mg·L−1。 -
电压的测量采用电压采集卡(PSIO813U,Honggeo.Ltd.Taiwan,China);盐度(以NaCl计)采用FE-30K型电导率仪测定;pH由PHS-3J酸度计测定;DO由WTW Oxi 7310溶解氧仪测定。COD、
${\rm{NH}}_4^ + $ -N、${\rm{NO}}_2^ - $ 、${\rm{NO}}_3^ - $ 、TN的检测采用国标法,由HACHDR6000紫外分光光度仪测定;取样及测量方法为:缓慢的抽取约8 mL样品溶液于称量瓶中,测量样品溶液的COD、${\rm{NH}}_4^ + $ -N、${\rm{NO}}_2^ - $ 、${\rm{NO}}_3^ - $ 、TN、pH、DO,测量完成后将多余的样品溶液缓慢的注射回各极室内,每次消耗的溶液量大约为1 mL。功率密度曲线与极化曲线的测量参考文献中的方法[1]。实验结束后,将实验阴极膜以及接种泥取出,于 −80 ℃下保存,送检进行高通量测序。 -
输出电压由电压采集卡每隔1 min测量1次,MFC功率通过式(1)进行计算,电流通过式(2)进行计算,硝酸盐平均去除速率由式(3)进行计算。
式中:P为MFC的功率,W;I为电流,A;U为输出电压,V。
式中:I为电流,A;R为外电阻阻值,Ω。
式中:V为硝酸盐平均去除速率,mg·(L·d)−1;C为硝酸盐的峰值浓度,mg·L−1;t为硝酸盐达到峰值到完全耗尽的时间,d。
硝酸盐平均去除速率无法精确的表征MFC的电营养反硝化作用。这主要是由于硝酸盐的峰值浓度通常发生在饥荒期,而饥荒期是一个不稳定的时期,存在硝化与内碳源反硝化的现象[13]。在稳定期虽然也发生内部碳源反硝化,但影响很小[6],因此,可用稳定期硝酸盐去除速率表述电营养反硝化速率。稳定期硝酸盐去除速率[12]由式(4)计算。
式中:Cs为稳定期硝酸盐浓度,mg·L−1;k为稳定期硝酸盐去除速率,mg·(L·d)−1;ts为运行时间,d;b为系数。
1.1. 实验用水与试剂



1.2. 实验装置与启动方法






1.3. 检测指标及方法






1.4. 电化学分析方法
-
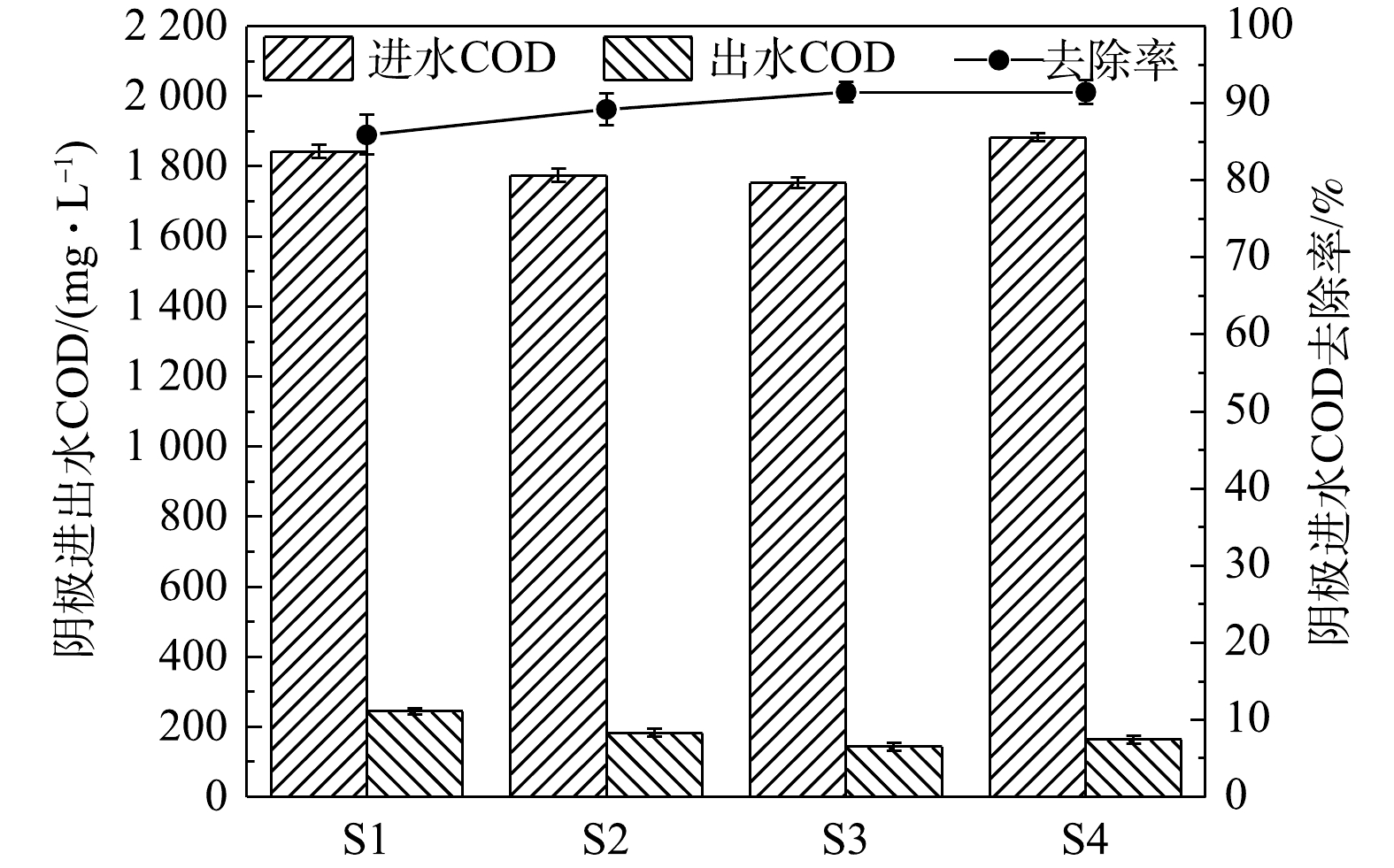
4组工况的阴极室进出水COD及去除率的变化如图3所示。其中S1、S2、S3、S4分别表示峰值电流为(0.24±0.03)、(0.37±0.03)、(0.55±0.11)、(0.5±0.2) mA时的工况。4组工况的阴极室进水COD去除率均在90%左右,其中S3实验组的COD去除率最高,达到了(91.44±1.29)%。相比较单室MFC[24-26],双室MFC的COD去除率同样较好。双室MFC是一种高效的有机物去除工艺,且可产生电能[27]。
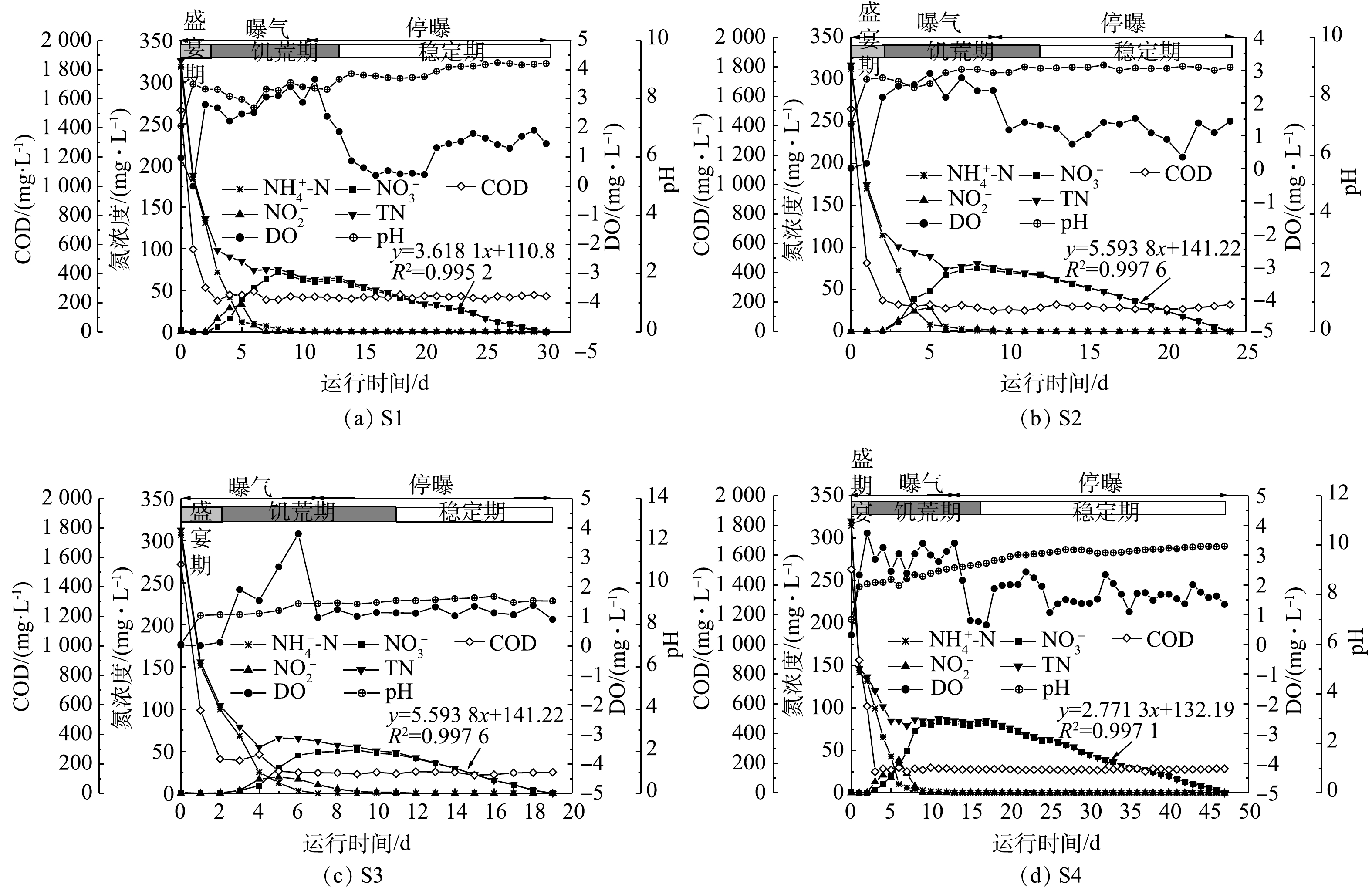
混合生物阴极MFC的1个运行周期可分为3个阶段:盛宴期,饥荒期与稳定期[12]。如图4所示,盛宴期主要特征为COD与
${\rm{NH}}_4^ + $ 的快速下降,但未检测出硝态氮,主要发生异养硝化/好氧反硝化反应;在饥荒期,由于阴极溶液中可被利用的耗氧有机物(以COD计)消耗殆尽,异养硝化/好氧反硝化反应随之减慢,自养硝化反应逐渐占主导地位,并伴随着一定的内碳源反硝化,此期间${\rm{NH}}_4^ + $ -N全部转化为硝酸盐并发生硝态氮的累积,TN的去除速率低;在稳定期,硝酸盐被电营养反硝化菌快速去除,且硝酸盐浓度的沿程变化呈线性下降趋势。盛宴期与饥荒期的区分指标为饥荒期开始出现${\rm{NO}}_2^ - $ 或${\rm{NO}}_3^ - $ ,而盛宴期的${\rm{NO}}_2^ - $ 或${\rm{NO}}_3^ - $ 浓度为0 mg·L−1。饥荒期与稳定期的区分方法为稳定期开始出现稳定的硝酸盐去除率,而饥荒期的硝酸盐去除率波动不定。停止曝气后DO基本维持在1~2 mg·L−1,溶解氧的偏高主要是由于阴极液的自然富氧作用[13]。另外,系统的pH对氮浓度的变化也有相应的响应。例如,阴极在${\rm{NH}}_4^ + $ 耗尽之前,整个系统的pH趋于降低,这是由于硝化作用引起的碱度消耗大于反硝化和氧还原过程中的碱度产生[13]。随着电化学反硝化作用和氧还原的进一步增强,系统的pH有所升高。阳极室
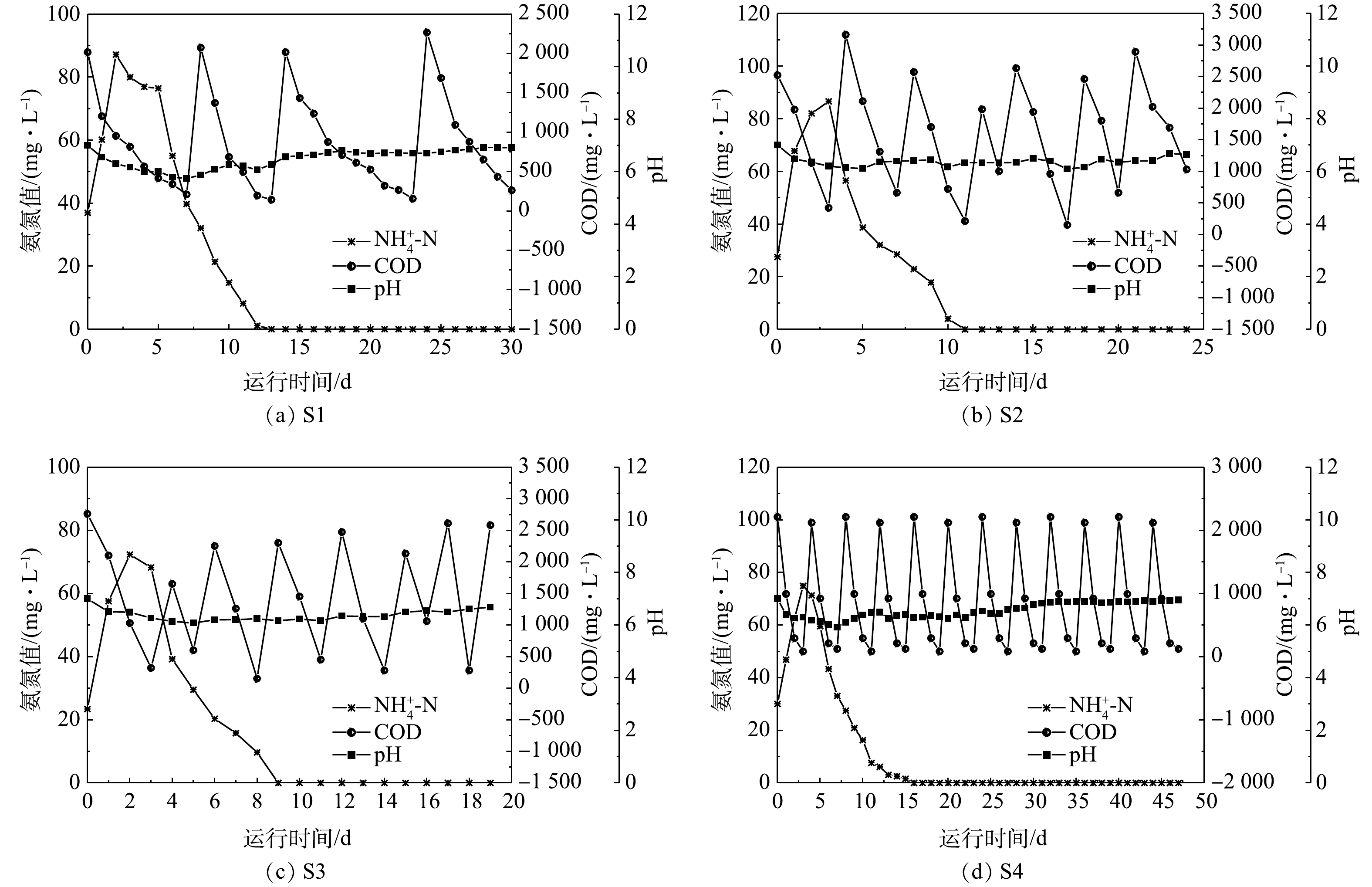
${\rm{NH}}_4^ + $ -N、COD和pH的沿程变化如图5所示。由于阳极室COD采用的投加方式为阶段补充,其变化曲线成锯齿形,控制其峰值大约在2 000 mg·L−1。阳极室的${\rm{NH}}_4^ + $ -N大致的趋势为先上升后下降,这是由于阳极室${\rm{NH}}_4^ + $ 的增减主要是通过离子扩散作用,而非阳极微生物的降解作用[26]。初期阳极室的${\rm{NH}}_4^ + $ 较阴极室低,阴极室的${\rm{NH}}_4^ + $ 向阳极室迁移,所以,初期的氨氮呈上升趋势;而后期随着阴极室${\rm{NH}}_4^ + $ 的不断减少,阳极室${\rm{NH}}_4^ + $ 低于阴极室,故向阴极室迁移[13]。因此,阳极室的氨氮去除速率亦能反映阴极微生物的硝化能力,并且由于阳离子交换膜的分隔,阳极室的${\rm{NH}}_4^ + $ -N去除具有滞后性。在本研究中,S3阳极室和阴极室的氨氮完全去除时间(9 d和7 d)最短,而其余3组分别为S1 (13 d和11 d)、S2 (11 d和9 d)和S4 (15 d和13 d)。4组工况在3个阶段的周期与氮去除率情况见表2。电流强度对盛宴期周期长短没有显著影响,均为(2±1) d,这可能是由于盛宴期周期很短。电流强度对MFC的稳定期周期影响最大,最大的稳定期周期为(30±1) d,而最小的稳定期周期为(7±1) d,变化幅度较大,且大致呈现出电流强度越大,稳定期周期越短的趋势。值得注意的是,电流强度对饥荒期周期长短也有影响,这说明随着硝酸盐的产生,生物电营养反硝化反应在饥荒期也扮演了一个不可忽略的角色。在盛宴期,4组反应器的
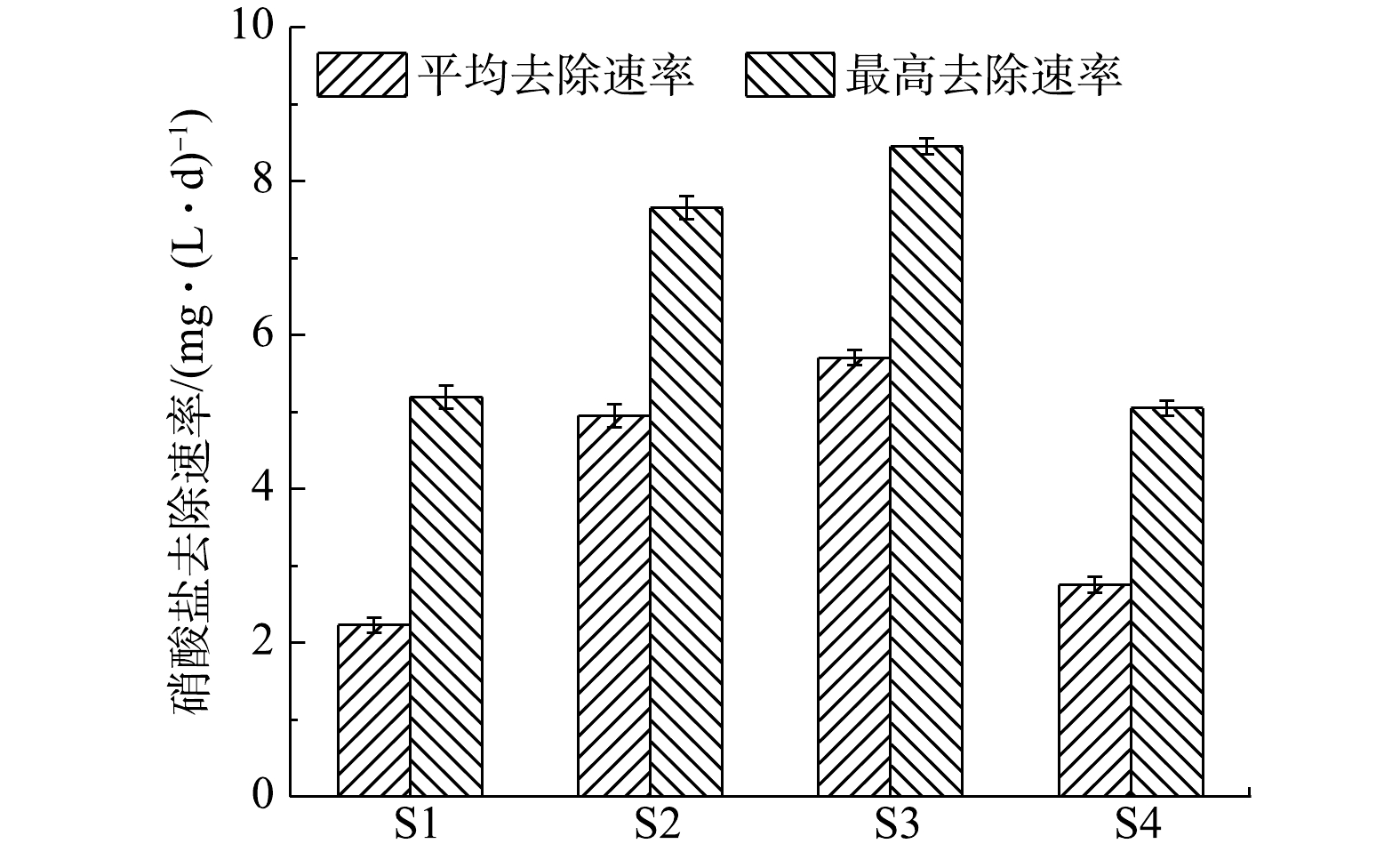
${\rm{NH}}_4^ + $ -N去除率(58.91%~67.61%)与TN去除率(58.14%~67.55%)均很高;在饥荒期,随着耗氧有机物(以COD计)的耗尽,氨氧化反应持续进行,但TN的去除率大幅度减慢,4组反应器的${\rm{NH}}_4^ + $ -N去除率为32.39%~41.82%,TN去除率在15.43%~21.45%;在稳定期,${\rm{NO}}_3^ - $ 被还原成N2,最后实现完全脱氮。与S1 (3.618 1 mg·(L·d)−1)、S2 (5.593 8 mg·(L·d)−1)、S4 (2.771 3 mg·(L·d)−1)相比,S3稳定期硝酸盐去除速率k (6.122 5 mg·(L·d)−1)最大,说明电流强度的增大有利于电营养反硝化反应进行,大的电子通量有利于电营养反硝化菌的生存。值得注意的是,相对于其他3组,S4的停曝期峰值电流(0.5±0.2 mA)较大,但稳定期硝酸盐去除速率并不高,为2.771 3 mg·(L·d)−1。这可能是由于S4的过低阴极电势(开路阴极电势为−110 mV)所造成的。在电营养反硝化系统中,阴极电势与电流强度同样重要[9,28]。与之前的研究类似,过低的阴极电势会导致硝酸盐的去除速率变低[12]。POUS等[28]研究了宽范围阴极电势(−703~597 mV)对BES中${\rm{NO}}_3^ - $ 去除速率的影响,发现阴极电势由597 mV降低至−403 mV的过程中,${\rm{NO}}_3^ - $ 的去除速率增加,但当阴极电势低于−403 mV时,${\rm{NO}}_3^ - $ 的去除速率却趋于平稳。另外,外电阻的减小,电子主要被非反硝化菌与氧气所消耗,也会导致系统的硝酸盐去除速率不佳。如图6所示,S1、S2、S3、S4实验组的硝酸盐平均去除速率分别达到(2.34±0.10)、(4.96±0.15)、(5.72±0.10)、(2.76±0.10) mg·(L·d)−1;硝酸盐最高去除速率分别为(5.18±0.15)、(7.65±0.13)、(8.45±0.15)、(5.05±0.11) mg·(L·d)−1。在4组实验中,S3实验组的硝酸盐平均去除速率与硝酸盐最高去除速率相对最高,这说明电流强度越大,反硝化速率越快。
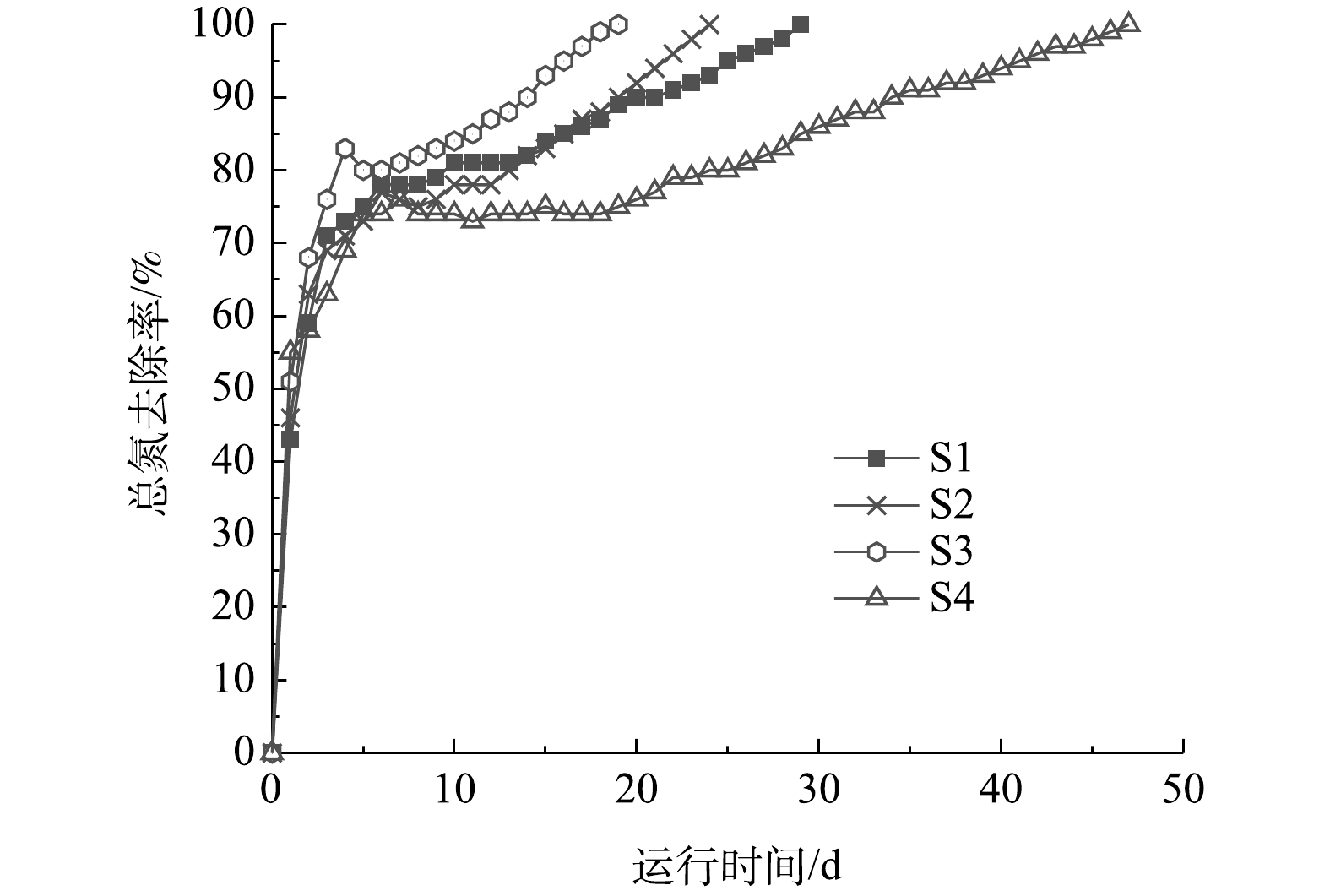
阴极室4组工况的总氮去除率如图7所示。总氮去除速率变化趋势为先增大后减少,这主要是由于盛宴期占主导优势的异养硝化/好氧反硝化脱氮速率大于饥荒期与稳定期的自养硝化与电营养反硝化[13]。与传统好氧/厌氧脱氮工艺相比较,异养硝化/好氧反硝化的TN去除速率更高[16]。本实验的总氮去除速率最高为7.20 mg·(L·d)−1。S1、S2、S3、S4的总氮去除率基本均在第5天达到最高值,为70%左右,且分别在第30、24、19、48天时实现完全脱氮。本实验最佳工况中总氮的去除负荷是0.111 kg·(m3·d)−1,而生物电化学的脱氮系统中的总氮的去除负荷通常是0.1~0.4 kg·(m3·d)−1[29-30],基本与本研究的结果相符。
-
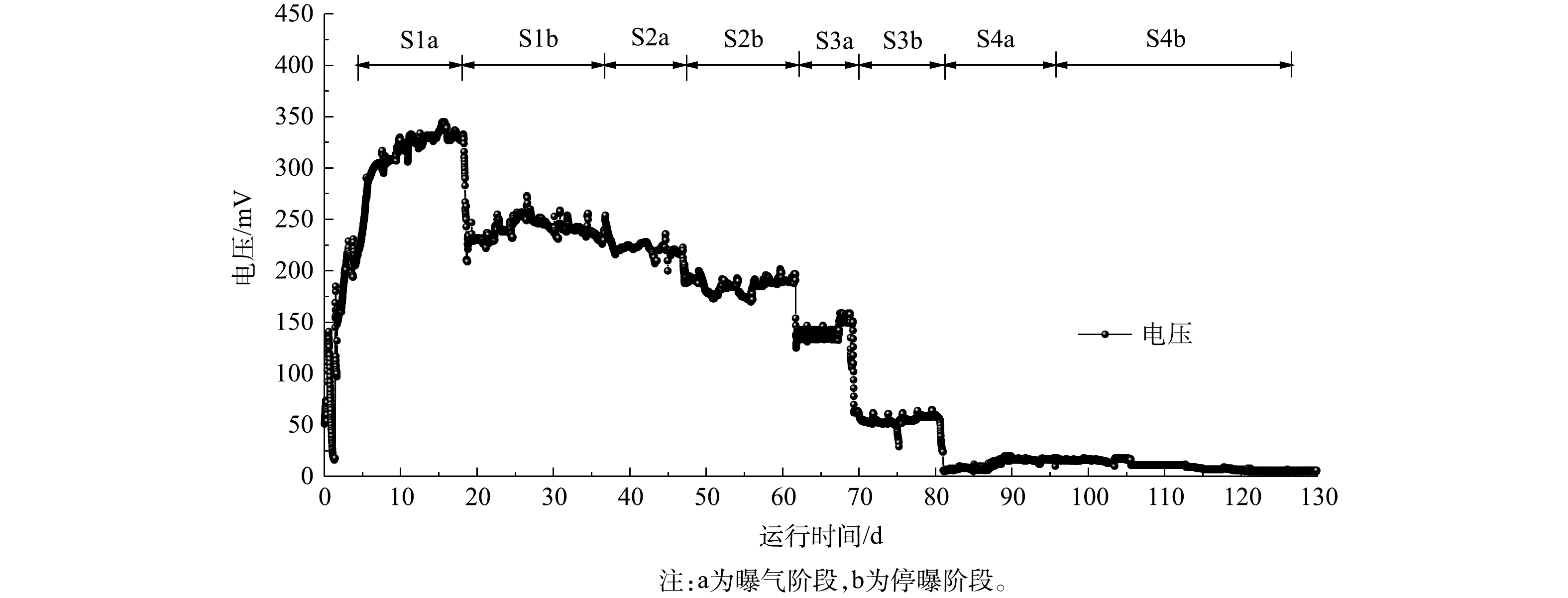
如图8所示,S1、S2、S3、S4曝气阶段峰值输出电压分别为(329±17)、(221±16)、(139±19)、(17±4) mV,由欧姆定律可知,与图8中上述数据对应的峰值电流分别为(0.33±0.01)、(0.44±0.03)、(1.39±0.19)、(0.85±0.2) mA;S1、S2、S3、S4停曝气阶段的峰值输出电压分别为(240±34)、(186±16)、(55±11)、(14±4) mV,同样上述数据对应的峰值电流分别是(0.24±0.03)、(0.37±0.03)、(0.55±0.11)、(0.5±0.2) mA。4组工况曝气状况下的电压与电流均大于非曝气状况。首先,停曝气阶段的阴极室溶解氧骤减,氧还原反应的减少将导致阴极电势与电流的降低[31]。其次,主要电子受体从氧气到硝酸盐的转变也是停曝阶段电流与电压降低的原因[32]。溶解氧的升高会带来输出电压的升高,系统的产电性能较好[33]。除了S1产电周期在144 h左右,S2,S3,S4产电周期均在72 h左右。
在系统运行的过程中,输出电压出现了不太稳定的情况。首先,当加入新鲜的阴极电解液时,普通好氧异养菌氧化有机物迅速增殖,好氧异养生物的竞争能力超过了阴极亲氧细菌或自养生物,对阴极电势和电池电压造成不利影响[34-35]。其次,生物硝化反应会消耗阴极室中的DO,阴极室电子受体的减少导致MFC产电性能的降低[32,36]。一旦有机物和
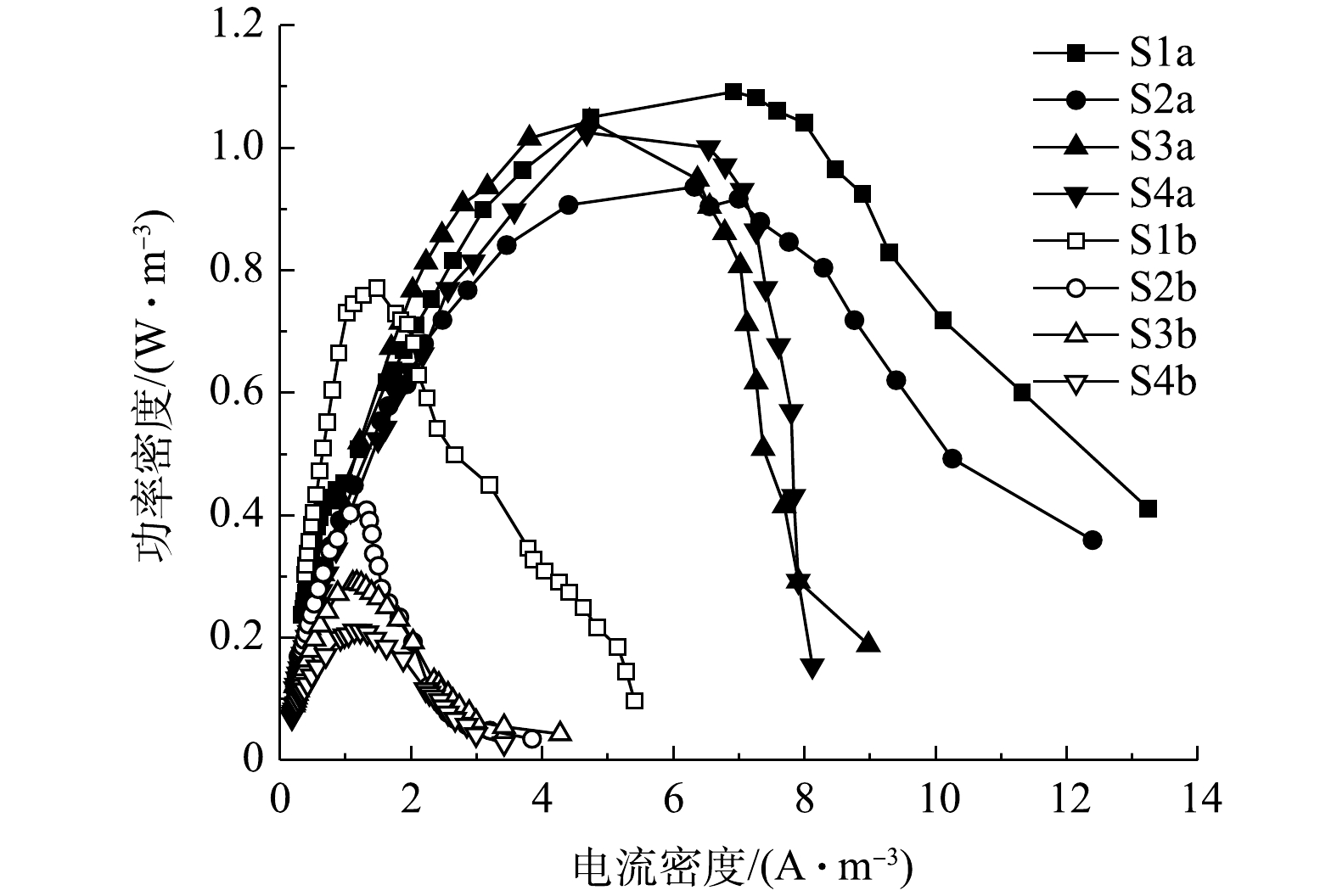
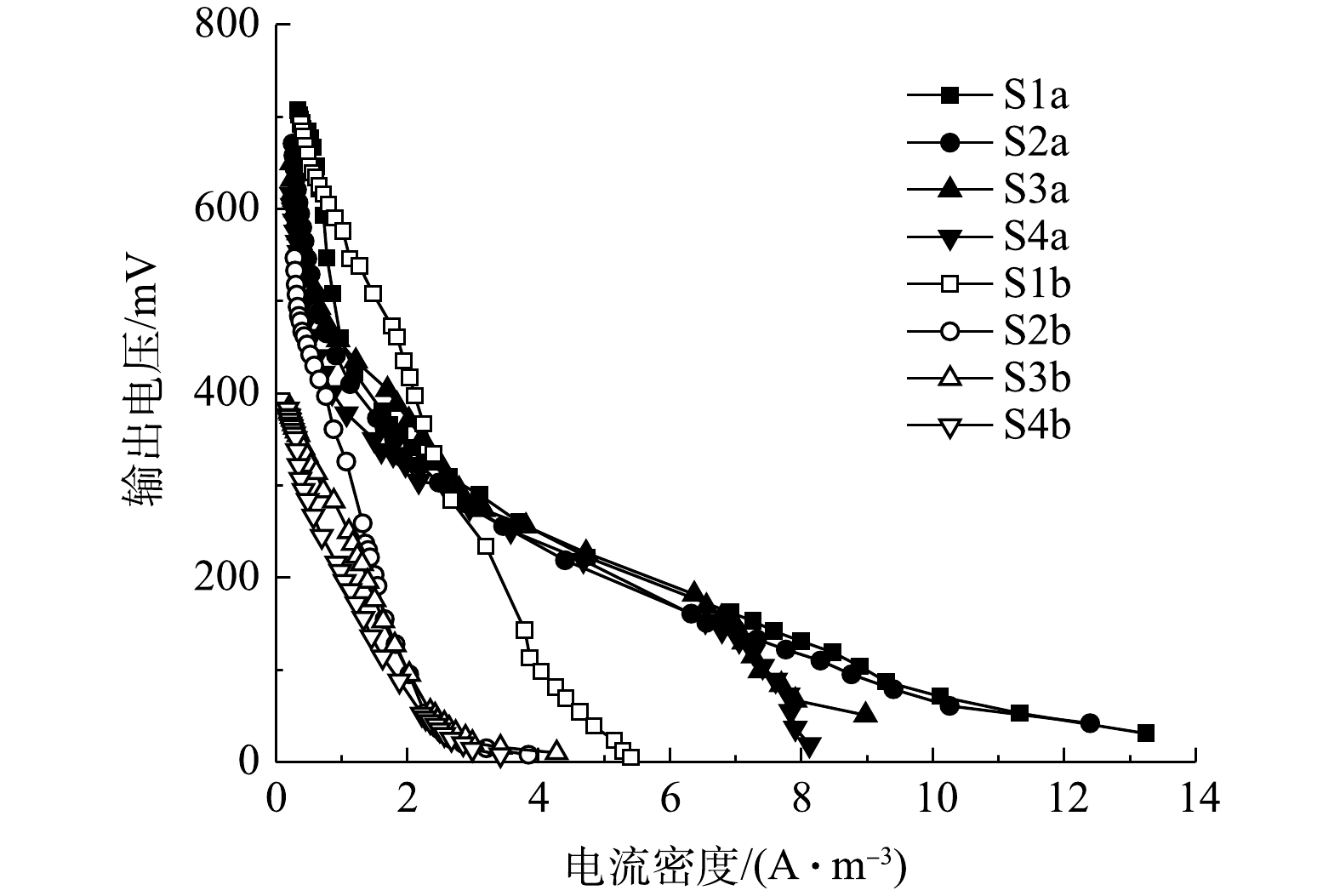
${\rm{NH}}_4^ + $ 耗尽,输出电压将逐渐上升并最终稳定。4组工况曝气与停曝阶段的极化曲线如图9和图10所示。由图9和图10中的数据可求得4组工况曝气与停曝阶段的MFC开路电压、最大功率密度及内部电阻,结果如表3所示。停曝阶段的最大功率密度变化趋势同开路电压变化趋势基本一致,且有随外电阻减小而减小的趋势,而曝气阶段的MFC性能差异并不显著。在混合生物阴极MFC的运行过程中,最大功率密度由实验开始的1.09 W·m−3减少至结束的0.21 W·m−3,开路电压也减少至412 mV,均有较大幅度地下降。造成MFC产电性能减少的原因有2个方面:第一,随着外部电阻的减小,由外部电阻获得的电压也越小[20],MFC的产电性能下降;第二,MFC长时间运行会加剧某些副反应的产生[37],如阳极的产甲烷反应[38]、质子交换膜污染[39]会降低MFC的输出功率与开路电压。
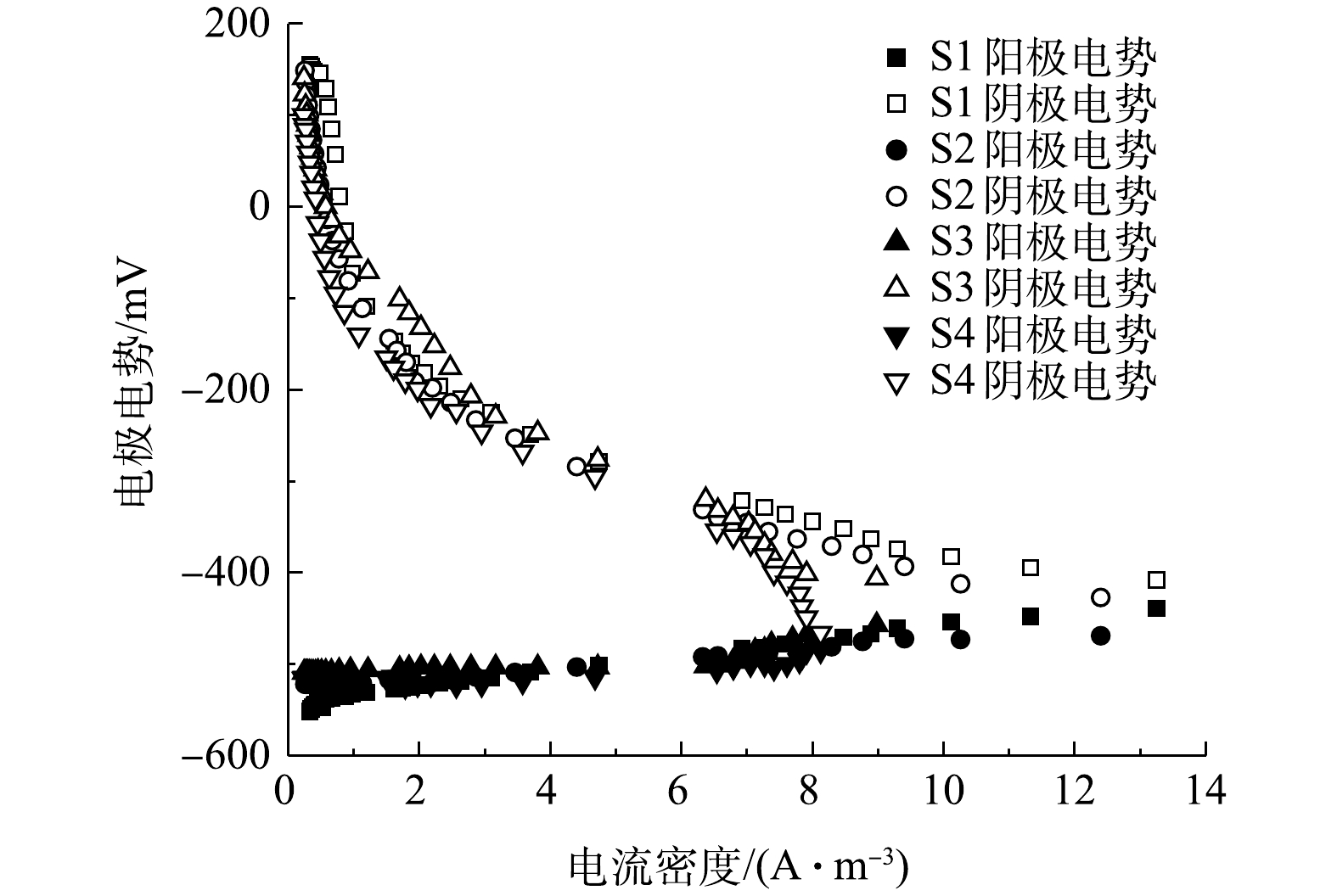
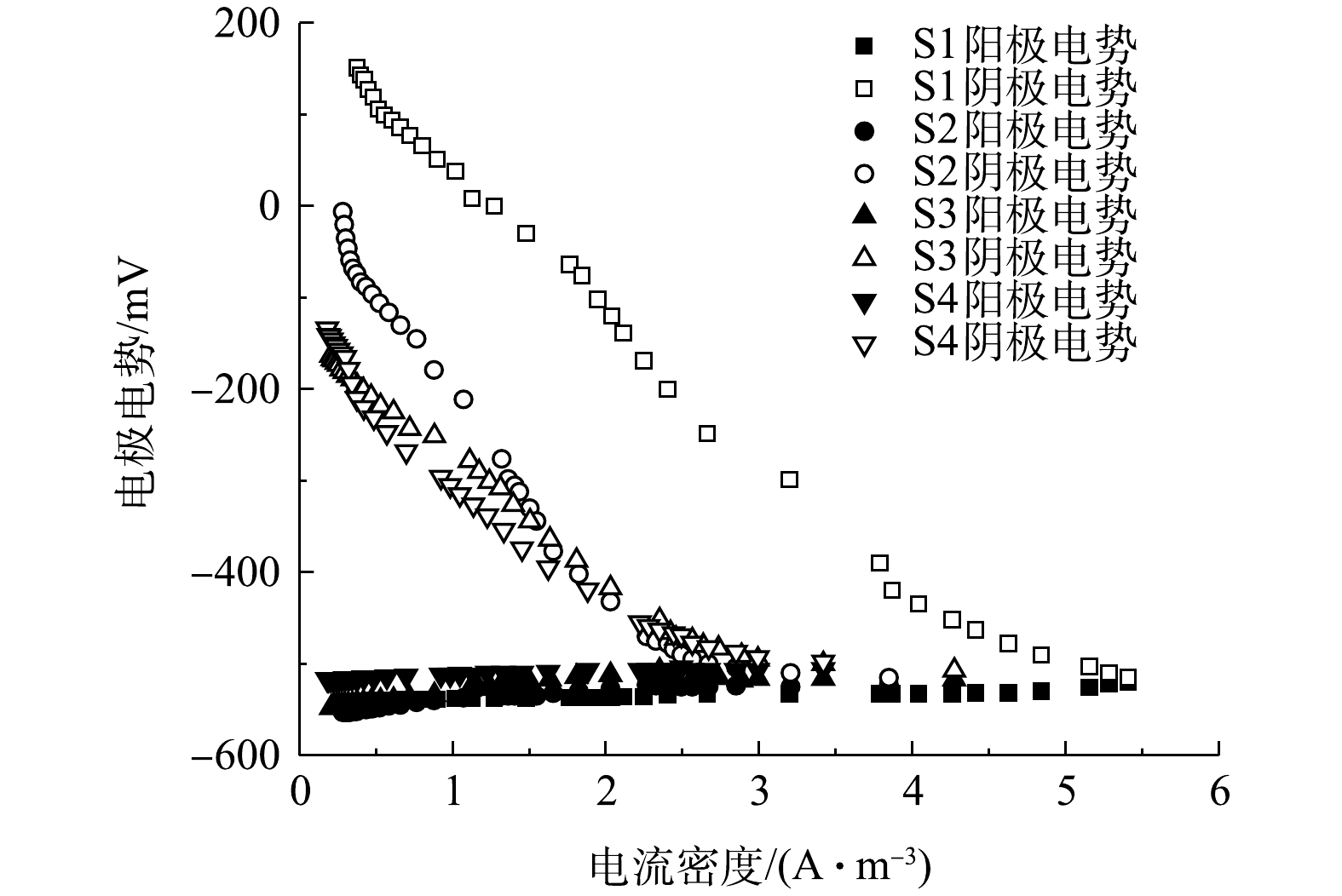
如图11和图12所示,在开路状况下,曝气阶段S1、S2、S3、S4的阴极电位分别为186,184,180,175 mV,阳极电位分别为−564、−545、−541、−524 mV,开路电压分别为750、729、721、699 mV;停曝气阶段S1、S2、S3、S4的阴极电位分别为186、40、−90、−110 mV,阳极电位分别为−560、−557、−552、−522 mV,开路电压分别为746、597、462、412 mV。曝气阶段的阴极电势普遍高于停曝阶段,这主要是由于高浓度的DO电子受体较多,导致较高的阴极电势,且DO越高,阴极过电势越低 [10]。与先前的研究类似[12],曝气与停曝阶段的阳极电势非常稳定。而阴极电势随着外电阻的增加,呈现出增大的趋势。
-
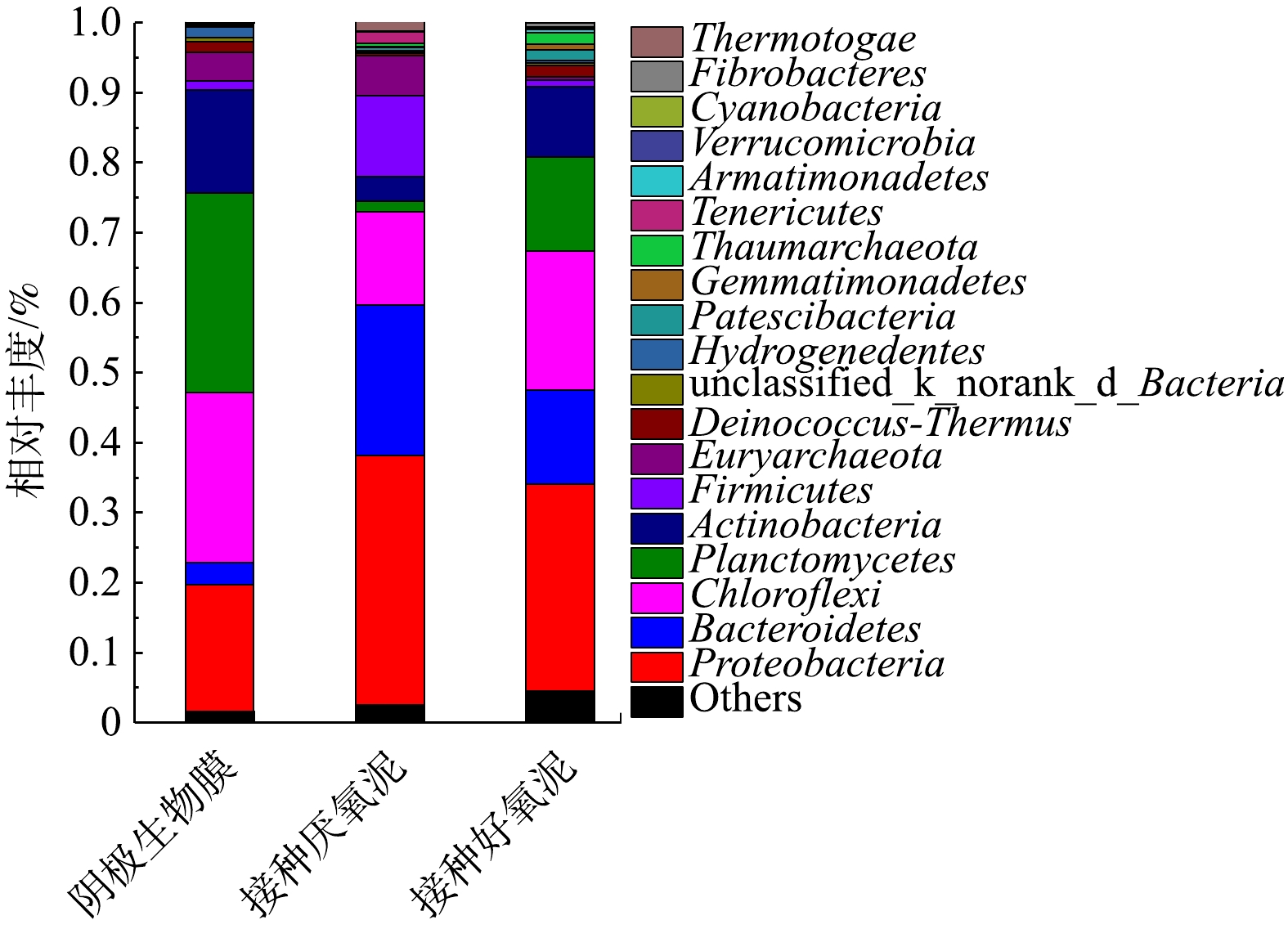
图13为门水平下阴极生物膜、接种厌氧泥与接种好氧泥的微生物群落分析。在门水平下,阴极生物膜相对丰度大于1% 的优势菌种为Planctomycetes (28.53%)、Chloroflexi (24.25%)、Proteobacteria (18.17%)、Actinobacteria (14.75%)、Euryarchaeota (4.06%)、Bacteroidetes (3.21%)、Deinococcus-Thermus (1.57%)、Hydrogenedentes (1.51%)、Firmicutes (1.24%)。接种厌氧泥的优势菌种为Proteobacteria (35.58%)、Bacteroidetes (21.43%)、Chloroflexi (13.42%)、Firmicutes (11.63%)、Euryarchaeota (5.62%)、Actinobacteria (3.52%)、Tenericutes (1.58%)、Planctomycetes (1.45%)、Thermotogae (1.20%)。接种好氧泥的优势菌种为Proteobacteria (29.56%)、Chloroflexi (19.79%)、Planctomycetes (13.50%)、Bacteroidetes (13.40%)、Actinobacteria (9.98%)、Deinococcus-Thermus (1.65%)、Thaumarchaeota (1.60%)、Patescibacteria (1.41%)。与接种污泥相比较,经MFC驯化后的生物阴极菌(门)落结构发生了改变,如生物阴极最大优势菌群为Planctomycetes,而接种厌氧泥与接种好氧泥则均为Proteobacteria。接种好氧泥与接种厌氧泥丰度均相对较高的Bacteroidetes,在阴极生物膜中仅占3.21%。Planctomycetes广泛的存于MFC阴极生物膜中,对生物阴极脱氮起着十分重要的作用[40]。ZHANG等[12]和WRIGHTON等[41]的研究表明,Proteobacteria是一类典型的高盐废水脱氮菌(门)落。Proteobacteria、Bacteroidetes是2种广泛存在的电化学活性菌(门),可较大幅度的提升MFC的产电性能[12,42]。
为了进一步剖析微生物的具体功能,对属水平下阴极生物膜、接种厌氧泥与接种好氧泥的微生物群落组成进行了更深一步阐述。如图14所示,阴极接种物和阴极生物膜之间在功能细菌种类在发生了巨大的变化。阴极膜与好氧接种泥相比,主要的AOB和NOB展现出差异性。好氧接种泥中主要的AOB为Nitrosomonas(1.78%)[43],阴极膜中仅含0.05%;好氧接种泥中主要的NOB为Nitrospira(1.47%),阴极生物膜中仅含0.06%。阴极生物膜中主要的AOB为SM1A02(22.12%),好氧接种泥仅含0.34%;阴极膜中主要的NOB为unclassified_f__Nitrosococcaceae (1.02%),好氧接种泥中仅含0.50%。厌氧接种泥主要的反硝化菌为Thauera (25.72%),而在阴极膜中仅为0.17%。阴极膜主要的反硝化菌为unclassified_f_Rhodobacteraceae(2.70%)、norank_f_Hydrogenedensaceae(1.5%)、unclassified_f_Anaerolineaceae (1.4%)、Oceanimonas (1.1%)、Azoarcus (0.7%)[30,44]。Nitrospira在好氧的环境下将
${\rm{NO}}_2^ - $ 氧化成${\rm{NO}}_3^ - $ [45]。Thauera是一类万能菌,可实现异养硝化/好氧反硝化,也是一种电营养反硝化菌[46-47]。Oceanimonas常出现在高盐废水处理的生物阴极中,可在厌氧的条件下将${\rm{NO}}_3^ - $ 还原成N2[48]。unclassified_f__Rhodobacteraceae (2.70%)具有异养硝化/好氧反硝化的功能[45],能将垃圾渗滤液中的
${\rm{NH}}_4^ + $ -N转化为${\rm{NO}}_2^ - $ 和${\rm{NO}}_3^ - $ ,又可以在曝气的条件下实现${\rm{NO}}_3^ - $ 的去除[15,44]。与其他研究异养硝化/好氧反硝化菌的相对丰度(5.06%)比相比较[13],本研究(2.70%)较低,这可能是由于S4实验组的“饥饿”时间较长导致的,长时间无碳源不利于异养硝化/好氧反硝化菌的生长[15]。SM1A02经常出现在高盐废水MFC的阴极生物膜检测结果中,是一类典型的高盐废水AOB[13]。unclassified_f__Nitrosococcaceae可将亚硝酸盐转化为硝酸盐[43]。生物阴极膜中承担起自养硝化功能的菌属有SM1A02、unclassified_f__Nitrosococcaceae。Hydrogenophaga和Azoarcus可利用无机物作为电子供体发生反硝化反应[49-50]。Hydrogenophaga已被证明是一类典型的电活性菌属,可提升MFC的产电性能[48]。值得注意的是,Azoarcu以被证明具有电营养反硝化功能[13]。生物阴极膜中可能承担着电营养反硝化功能的菌属为Hydrogenophaga和Azoarcus。
2.1. 污染物去除情况





















2.2. 产电性能分析

2.3. 微生物群落分析







-
1)高电流强度可缩短双室混合生物阴极MFC的完全脱氮周期,且主要缩短混合生物阴极MFC的稳定期周期。在4组实验中,当电流强度((0.55±0.11) mA)最大时,硝酸盐去除速率最大,实现总氮完全去除的时间(19 d)最短。
2)电流强度的增大有利于混合生物阴极MFC电营养反硝化反应的进行,但过小的外接电阻会造成阴极电势的降低,从而降低硝酸盐去除速率。
3)在曝气阶段,混合生物阴极MFC系统外加电阻越大,电池的开路电压亦越大,而最大功率密度却相差不大;在停止曝气阶段,由于阴极室电子受体的减少,MFC产电性能普遍下降,且系统的开路电压与最大功率密度均随着外加电阻的增大而增大。
4)高通量测序的结果表明,承担起氮的去除功能的相关微生物有SM1A02、unclassified_f__Rhodobacteraceae、unclassified_f__Nitrosococcaceae、norank_f_Hydrogenedensaceae、unclassified_f_Anaerolineaceae、Oceanimonas、Azoarcus、unclassified_f_Rhodobacteraceae。其中norank_f_Hydrogenophaga和Azoarcus可能承担电营养反硝化功能。
